



Introduction
Lung cancer has become one of the leading causes of mortality in the world [1]. Of its different pathological subtypes, non-small cell lung cancer (NSCLC) accounts for about 85% of all lung cancer cases, and lung adenocarcinoma (LUAD) constitutes the highest proportion of NSCLC cases [2]. The relationship between the expression of certain genes and LUAD progression has been increasingly investigated [3–5]. For example, the decreased expression of TNFAIP3, an anti-inflammatory gene that helps maintain the inflammatory balance, reduced the proportion of CD8+ T cells, thus assisting LUAD cells in immune evasion. Moreover, it caused anti-programmed cell death protein 1 monotherapy to be ineffective and led to poor prognosis in LUAD patients [5].
The expression level of SLC7A8, which encodes large neutral amino acid transporter small subunit 2, was shown to be related to tumor progression [6–9]. SLC7A8 was highly expressed in estrogen receptor (ER)-positive breast cancer and was a marker of good prognosis in these patients [6]. In uterine leiomyoma tissues, luteinizing hormone upregulated SLC7A8. Knockdown of SLC7A8 enhanced the proliferation of uterine leiomyoma cells [7]. A mutation in SLC7A8 reduced the efficacy of the combination of cisplatin and verapamil for treating esophageal squamous carcinoma and induced the development of drug resistance [8]. Asada et al. showed that SLC7A8 overexpression improved the survival rate of LUAD patients [9]. However, the role of SLC7A8 in LUAD progression is still obscure.
This study aimed to further elucidate the relationship between SLC7A8 expression and LUAD prognostic factors using bioinformatics and cell models. We also examined the role of SLC7A8 in LUAD cell growth and metastasis in vitro.
Results
The expression of SLC7A8 was attenuated in LUAD
The transcripts per million (TPM) data retrieved from The Cancer Genome Atlas (TCGA) and Xena showed that SLC7A8 was downregulated in LUAD tissues compared with normal lung tissues (Figure 1). The expression of SLC7A8 was lower in unpaired LUAD tissues than in normal tissues from TCGA and Xena (Figure 1A, 1B) and in paired LUAD tissues from TCGA (Figure 1C).
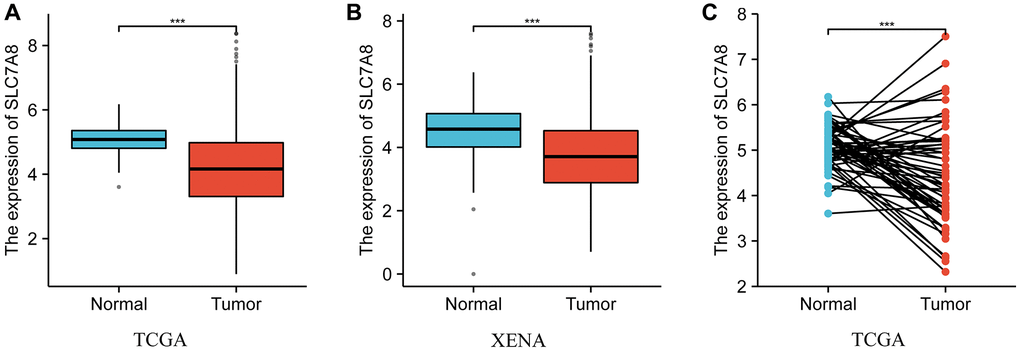
Figure 1. The expression level of SLC7A8 in LUAD based on TPM data from TCGA and Xena. (A) TPM data of unpaired tissues from TCGA. (B) TPM data of unpaired tissues from Xena. (C) TPM data of paired tissues from TCGA. Abbreviations: LUAD: lung adenocarcinoma; TCGA: The Cancer Genome Atlas; TPM: transcripts per million.
Diagnostic value of SLC7A8 in LUAD
Receiver operating characteristic (ROC) curve analysis indicated that the expression level of SLC7A8 carried significant diagnostic value in LUAD. The area under the curve values of ROC curves plotted based on SLC7A8 expression data from TCGA and Xena were 0.750 and 0.707, respectively (Figure 2A, 2B), highlighting the important diagnostic value of SLC7A8 in LUAD.
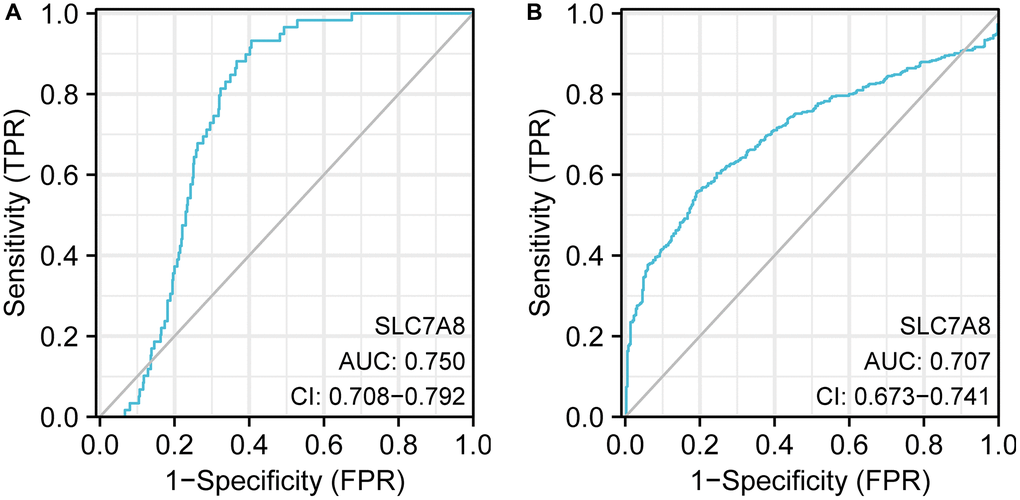
Figure 2. ROC curve analysis indicating the diagnostic value of SLC7A8 in LUAD. (A) TPM data from TCGA. (B) TPM data from Xena. Abbreviation: ROC: receiver operating characteristic.
Low expression of SLC7A8 was associated with poor prognosis in LUAD patients
Prognostic data based on the TPM values from TCGA revealed that low expression of SLC7A8 was associated with poor prognosis in LUAD patients, especially with shorter overall survival (OS), disease-specific survival (DSS), and progression-free interval (PFI) (Figure 3A–3C). Cox regression analysis (Table 1) confirmed the association between low SLC7A8 expression and poor patient prognosis. A nomogram constructed based on the primary therapy outcome, T stage, N stage, M stage, and SLC7A8 expression accurately predicted patient prognosis (Figure 4).
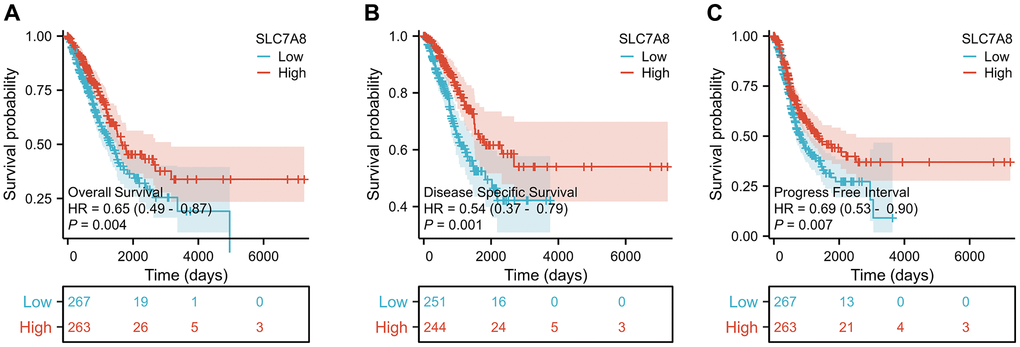
Figure 3. Expression level of SLC7A8 is associated with the prognosis of LUAD patients. SLC7A8 expression is associated with (A) OS in LUAD patients, (B) DSS in LUAD patients, (C) PFI in LUAD patients. Abbreviations: OS: overall survival; DSS: disease-specific survival; PFI: progression-free interval.
Table 1. Cox regression analysis reveals the prognostic factors in LUAD.
| Characteristics | N | HR (95% CI) | P-value |
| T stage | 527 | <0.001 | |
| T1 | 176 | Reference | |
| T2 | 285 | 1.507 (1.059–2.146) | 0.023 |
| T3 and T4 | 66 | 3.095 (1.967–4.868) | <0.001 |
| N stage | 514 | <0.001 | |
| N0 | 345 | Reference | |
| N1 | 96 | 2.293 (1.632–3.221) | <0.001 |
| N2 and N3 | 73 | 2.993 (2.057–4.354) | <0.001 |
| M stage | 381 | 0.010 | |
| M0 | 356 | Reference | |
| M1 | 25 | 2.176 (1.272–3.722) | 0.005 |
| Primary therapy outcome | 442 | <0.001 | |
| PD | 71 | Reference | |
| SD | 38 | 0.289 (0.141–0.592) | <0.001 |
| PR | 5 | 0.702 (0.170–2.897) | 0.625 |
| CR | 328 | 0.266 (0.185–0.382) | <0.001 |
| Race | 472 | 0.191 | |
| Asian | 8 | Reference | |
| Black or African American | 55 | 1.911 (0.254–14.382) | 0.529 |
| White | 409 | 2.714 (0.380–19.403) | 0.320 |
| Gender | 530 | 0.570 | |
| Female | 283 | Reference | |
| Male | 247 | 1.087 (0.816–1.448) | 0.569 |
| Age | 520 | 0.185 | |
| ≤65 | 257 | Reference | |
| >65 | 263 | 1.216 (0.910–1.625) | 0.186 |
| SLC7A8 | 530 | 0.004 | |
| Low | 267 | Reference | |
| High | 263 | 0.655 (0.491–0.874) | 0.004 |
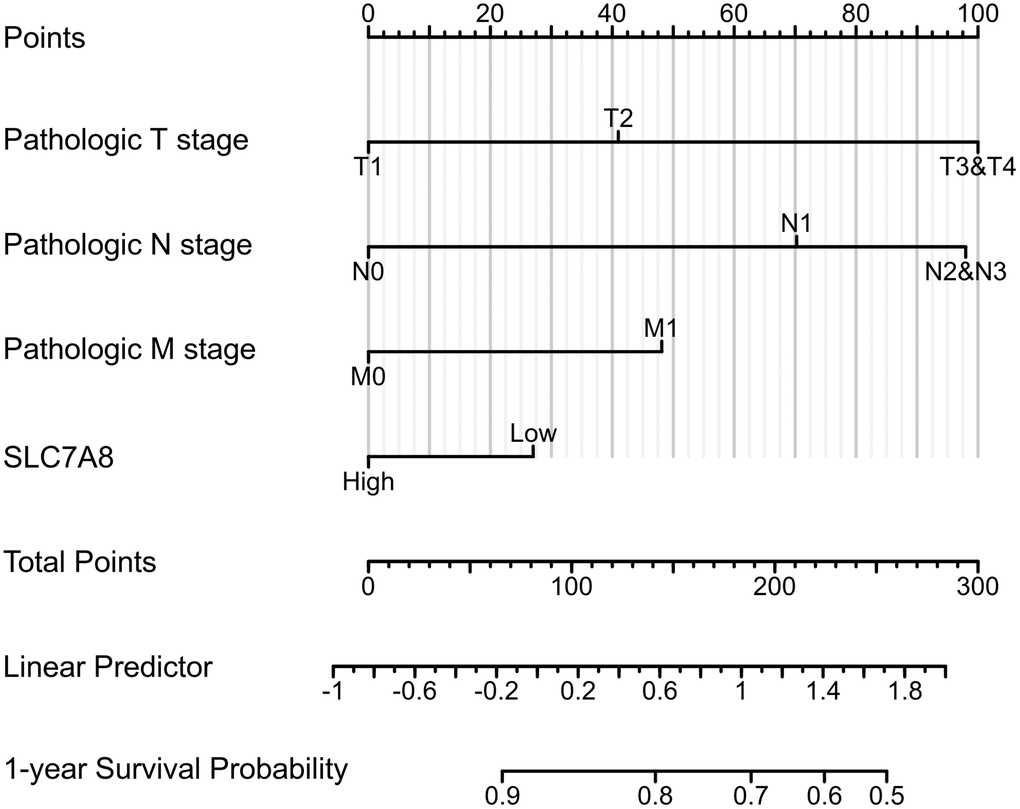
Figure 4. The nomogram shows the correlation between low expression of SLC7A8 and poor prognosis of LUAD.
SLC7A8 overexpression inhibited the proliferation, migration, and invasion of LUAD cells
The expression of SLC7A8 was significantly higher in BEAS-2B cells than in A549 and H1299 cells (Figure 5A). Gene Ontology analysis showed that 679 genes co-expressed with SLC7A8 were correlated with cell cycle progression (Table 2). The overexpression of SLC7A8 in A549 and H1299 cells was successfully verified using Western blotting (Figure 5B). The Transwell assay showed that the number of A549 and H1299 cells in the SLC7A8-overexpression group were significantly lower than in the control group (Figure 5C). Overexpressing SLC7A8 inhibited the migration of A549 and H1299 cells, as evidenced by the wound healing assay (Figure 5D). The Cell Counting Kit-8 (CCK-8) assay also demonstrated that the number of A549 and H1299 cells significantly decreased in the SLC7A8-overexpression group compared with the control group (Figure 6A). The colony forming assay showed that the ability of cells to proliferate was lower in the SLC7A8-overexpression group (Figure 6B). These findings indicate that overexpressing SLC7A8 inhibited the proliferation, migration, and invasion of LUAD cells.
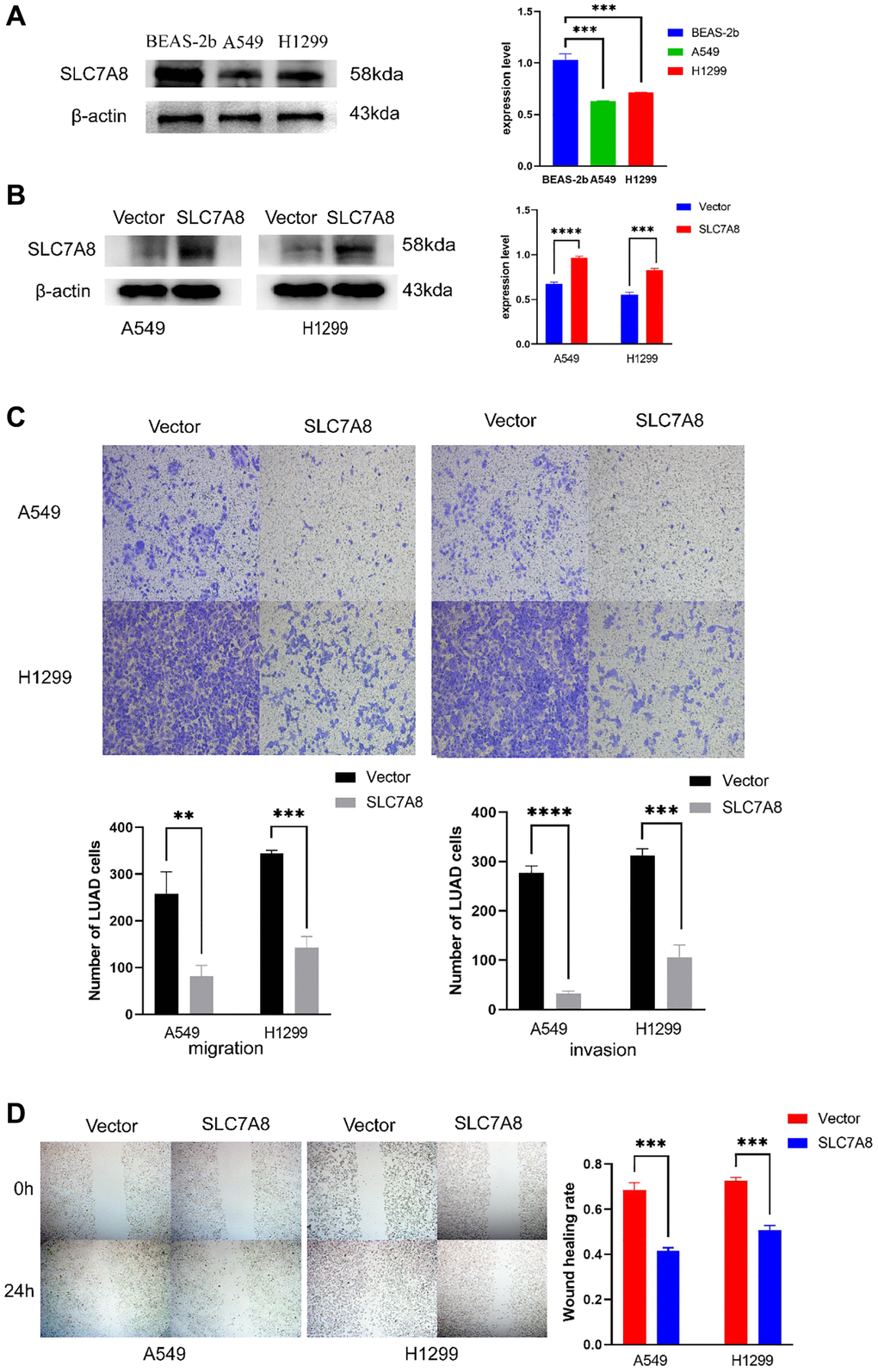
Figure 5. Overexpression of SLC7A8 suppresses the migration and invasion of LUAD. (A) The expression level of SLC7A8 in BEAS-2B, A549, and H1299 cells. (B) The protein expression levels were determined by Western blotting. (C) The migration and invasion were assessed using Transwell assays. (D) The migration ability was determined using wound healing assays as well.
Table 2. GO/KEGG analysis of SLC7A8.
| Type | ID | Description | P-value | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0018958 | phenol-containing compound metabolic process | 6.1974E-06 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0030038 | contractile actin filament bundle assembly | 1.73008E-05 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0043149 | stress fiber assembly | 1.73008E-05 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0016999 | antibiotic metabolic process | 3.44352E-05 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0006067 | ethanol metabolic process | 4.15677E-05 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0017000 | antibiotic biosynthetic process | 4.15677E-05 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0031638 | zymogen activation | 4.78141E-05 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0006687 | glycosphingolipid metabolic process | 5.08109E-05 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0006664 | glycolipid metabolic process | 5.9784E-05 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:1903509 | liposaccharide metabolic process | 6.57364E-05 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0031032 | actomyosin structure organization | 9.37131E-05 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0006643 | membrane lipid metabolic process | 0.000154598 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0050665 | hydrogen peroxide biosynthetic process | 0.000160988 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0072073 | kidney epithelium development | 0.00019478 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0045056 | transcytosis | 0.000216961 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0001655 | urogenital system development | 0.000220889 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0006665 | sphingolipid metabolic process | 0.000222584 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0072001 | renal system development | 0.000256287 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0061337 | cardiac conduction | 0.000301821 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0051492 | regulation of stress fiber assembly | 0.000393813 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0032355 | response to estradiol | 0.000438313 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0006068 | ethanol catabolic process | 0.000447416 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0072520 | seminiferous tubule development | 0.000447416 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0042403 | thyroid hormone metabolic process | 0.000475202 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0051017 | actin filament bundle assembly | 0.000487038 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0030324 | lung development | 0.000518227 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0071560 | cellular response to transforming growth factor beta stimulus | 0.000542241 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0042445 | hormone metabolic process | 0.000618613 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0061572 | actin filament bundle organization | 0.000630999 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0031670 | cellular response to nutrient | 0.000650715 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0051145 | smooth muscle cell differentiation | 0.000650715 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:1901616 | organic hydroxy compound catabolic process | 0.000650715 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0030323 | respiratory tube development | 0.000659255 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0007015 | actin filament organization | 0.000692946 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0071559 | response to transforming growth factor beta | 0.000724183 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0051056 | regulation of small GTPase mediated signal transduction | 0.000749109 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0060541 | respiratory system development | 0.000789527 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0006631 | fatty acid metabolic process | 0.000820985 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0021692 | cerebellar Purkinje cell layer morphogenesis | 0.000858959 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0097202 | activation of cysteine-type endopeptidase activity | 0.000858959 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BP | GO:0043268 | positive regulation of potassium ion transport | 0.000865894 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CC | GO:0042599 | lamellar body | 0.000150683 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CC | GO:0062023 | collagen-containing extracellular matrix | 0.000312972 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| KEGG | hsa00280 | Valine, leucine and isoleucine degradation | 4.78595E-06 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| KEGG | hsa00380 | Tryptophan metabolism | 1.1307E-05 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| KEGG | hsa00340 | Histidine metabolism | 8.15346E-05 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| KEGG | hsa05224 | Breast cancer | 0.000186964 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| KEGG | hsa00410 | beta-Alanine metabolism | 0.000512328 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| KEGG | hsa00071 | Fatty acid degradation | 0.000757955 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| KEGG | hsa00600 | Sphingolipid metabolism | 0.001463422 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| KEGG | hsa04142 | Lysosome | 0.00171453 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| KEGG | hsa05226 | Gastric cancer | 0.00214208 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| KEGG | hsa00620 | Pyruvate metabolism | 0.002161182 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| KEGG | hsa04115 | p53 signaling pathway | 0.003838081 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| KEGG | hsa04110 | Cell cycle | 0.004060117 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Abbreviations: GO: Gene Ontology; KEGG: Kyoto Encyclopedia of Genes and Genomes. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
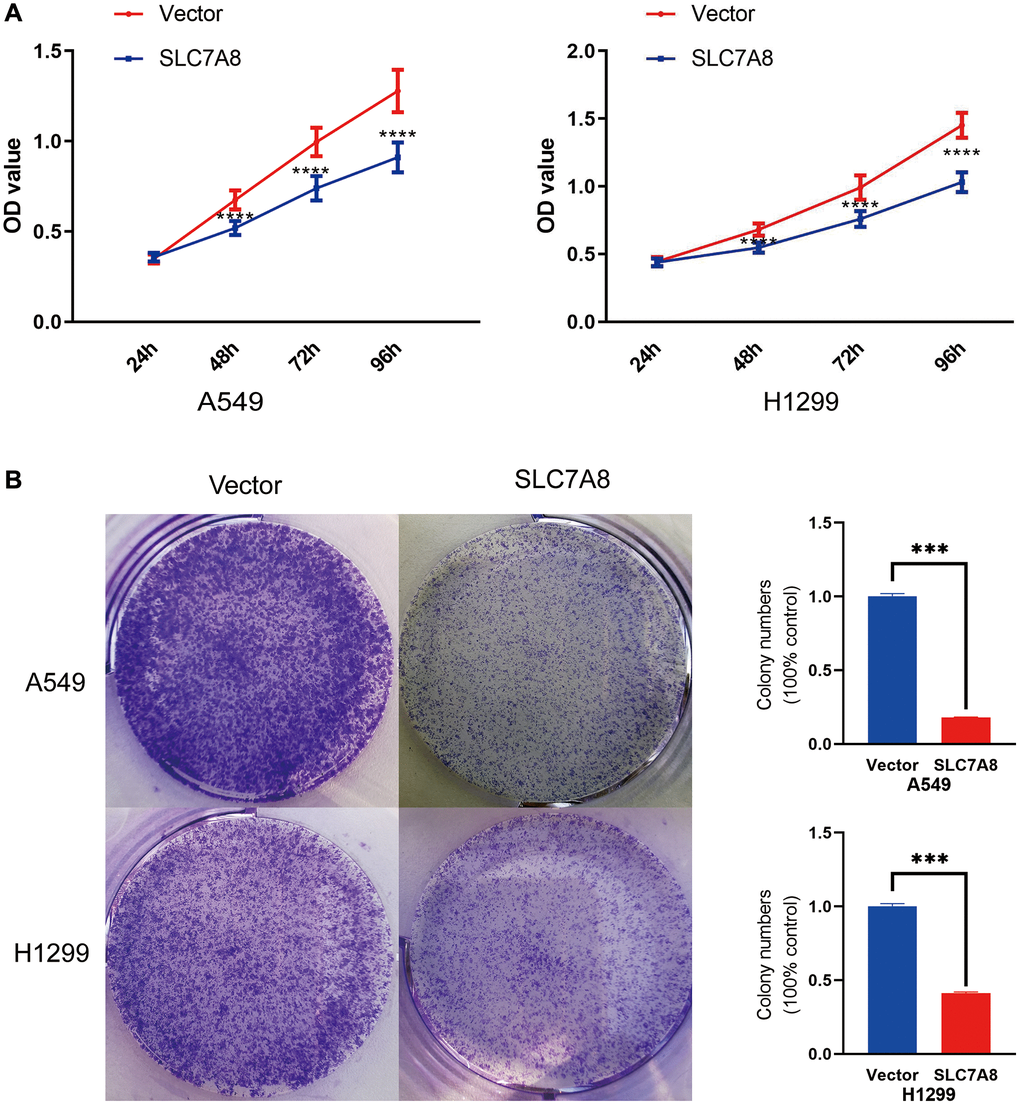
Figure 6. (A) Cell proliferation in the SLC7A8-overexpressing group was demonstrated using the CCK-8 assays. (B) Cell proliferation in the SLC7A8-overexpressing group was demonstrated using and colony forming assays. Abbreviation: CCK-8: Cell Counting Kit-8.
Mechanisms and protein–protein interaction (PPI) network of SLC7A8
The PPI network shows protein relationships among SLC7A8-related genes (Figure 7). Kyoto Encyclopedia of Genes and Genomes enrichment analysis and Gene Set Enrichment Analysis (GSEA) indicated that SLC7A8 influenced the cell cycle and DNA replication (Figure 8A–8F and Table 2). We conducted relevant experiments to validate these preliminary findings and found that the expression of cyclin-dependent kinase-1 (CDK1) and cyclin B1 (CCNB1), two important cell cycle proteins, was significantly attenuated in LUAD cells overexpressing SLC7A8 (Figure 8G). These results suggest that overexpressing SLC7A8 may potentially hinder the progression of LUAD by inhibiting the cell cycle pathway.
Figure 7. PPI network of genes strongly co-expressed with SLC7A8. Abbreviation: PPI: protein–protein interaction.
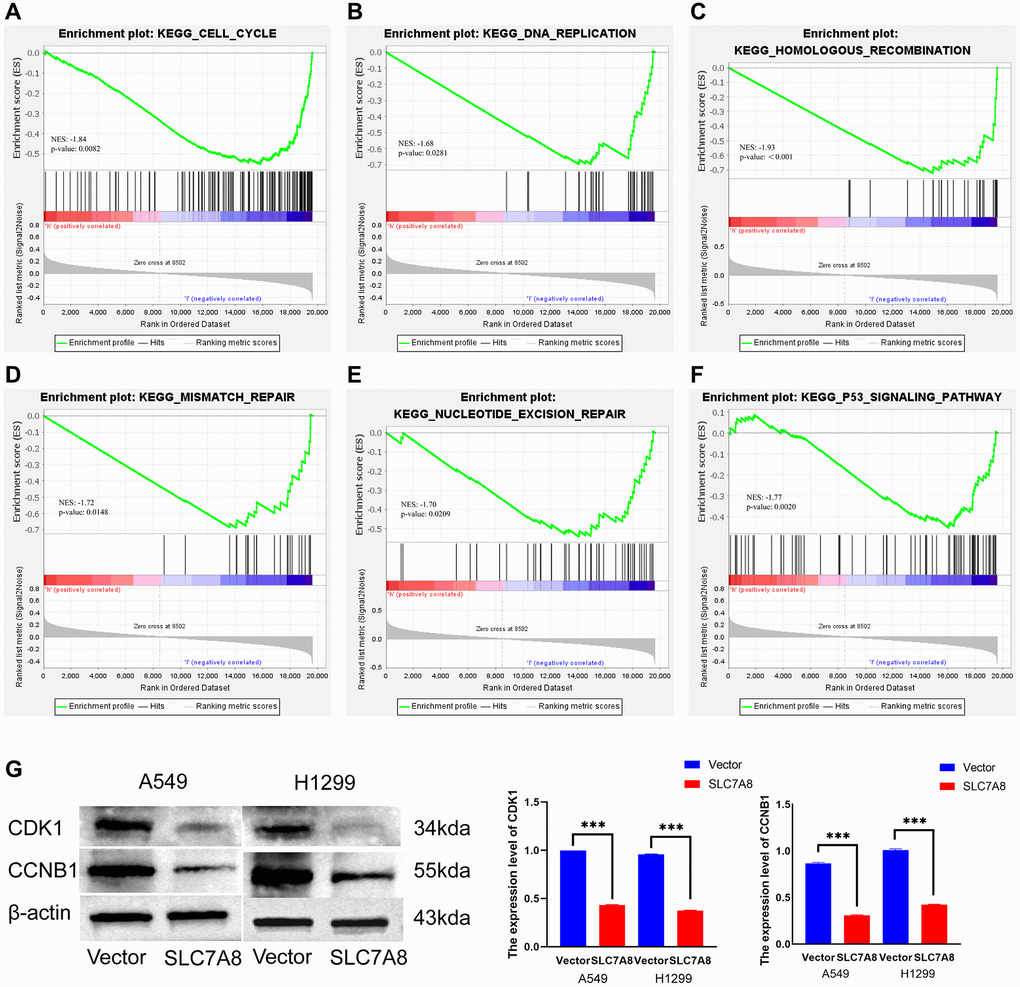
Figure 8. Mechanisms associated with SLC7A8. (A–F) Mechanisms associated with SLC7A8 were explored using GSEA. (G) The expression levels of the cell cycle proteins CDK1 and CCNB1. Abbreviations: GSEA: Gene Set Enrichment Analysis; CDK1: cyclin-dependent kinase-1; CCNB1: cyclin B1.
SLC7A8 expression was correlated with infiltrated immune cells and immune factors in LUAD
The expression level of SLC7A8 was significantly correlated with the level of immune cell infiltration in LUAD, especially that of B cells, interstitial dendritic cells, mast cells, CD56 bright cells, natural killer cells, plasmacytoid DCs, T follicular helper cells, T helper 2 cells, T helper17 cells (Figure 9). SLC7A8 expression was also associated with some immune factors, such as CD5, CD34, CD36, CD37, CD79B, CDA, CDC7, CCL4, and CCL28 (Figure 10).
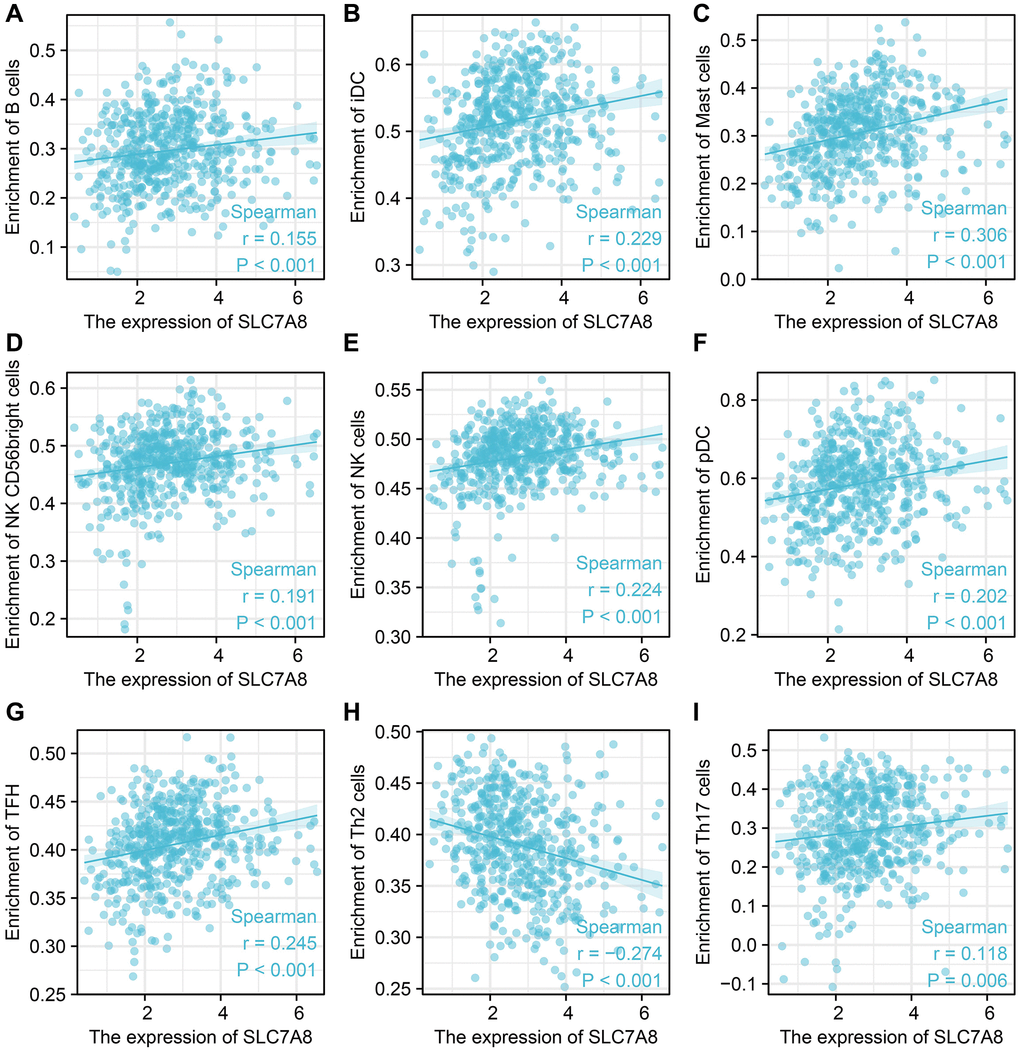
Figure 9. SLC7A8 expression is correlated with immune infiltrated cells. (A) B cells. (B) iDC. (C) Mast cells. (D) NK CD56 bright cells. (E) NK cells. (F) pDC. (G) TFH. (H) Th2 cells. (I) Th17 cells.
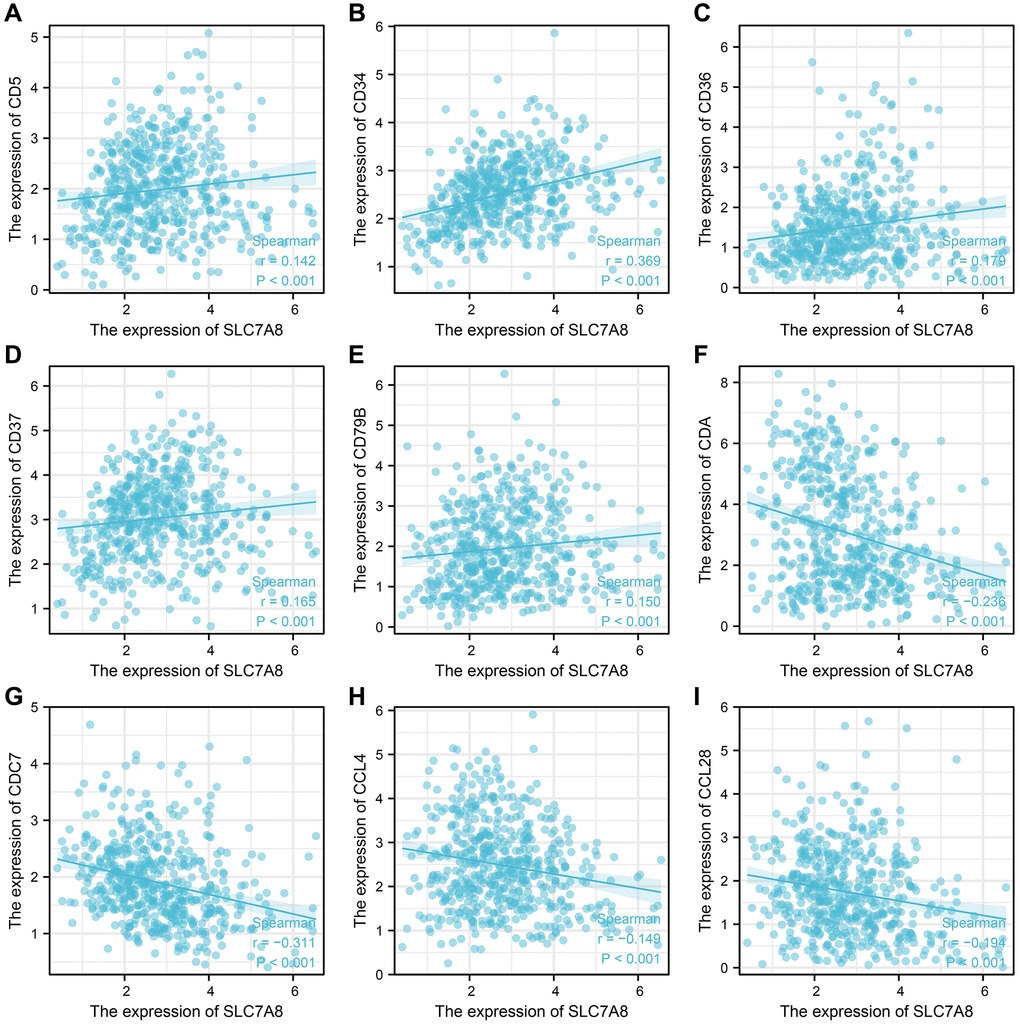
Figure 10. SLC7A8 expression is correlated with immune factors. (A) CD5. (B) CD34. (C) CD36. (D) CD37. (E) CD79B. (F) CDA. (G) CDC7. (H) CCL4. (I) CCL28.
Discussion
Although progress in the treatment and early screening of lung cancer have mitigated its incidence and mortality, it still accounts for a high percentage of cancer-related deaths [10]. Studies have identified some genes associated with cancer progression. SLC7 genes were reported to be correlated with the prognosis of lung cancer [11–14]. SLC7A5 was overexpressed in LUAD and significantly correlated with the Ki-67 labeling index, which can increase metabolic activity and is associated with tumor cell growth [11]. SLC7A8 was shown to be a good prognostic marker for breast cancer. When overexpressed, it behaved like a tumor suppressor, especially in low-growth ER-positive subtypes [6]. Jiang et al. found that SLC7A2 increased drug sensitivity, immune infiltration, and survival in NSCLC [12]. The upregulation of SLC7A7 increased the levels of arginine in tumor cells, thus promoting their migration and invasion [13]. Conversely, knockdown of SLC7A9 inhibited the migration, invasion, and proliferation of esophageal squamous cell carcinoma cells, making it a suitable biomarker to predict lymph node metastasis of this cancer [14]. Our studies showed that SLC7A8 was lowly expressed in LUAD, whereas overexpression of SLC7A8 was associated with higher OS, DSS, and PFI, making it a potential prognostic factor for LUAD.
The immune microenvironment intimately affects the progression of LUAD [15–17]. Cancer-related immune cells were shown to serve as biomarkers for the progression of NSCLC [18]. Genes from the SLC7 family, such as SLC7A11, were shown to be associated with the immune microenvironment [19]. B cells, which are involved in humoral immunity, were shown to be an important component of the immune microenvironment of lung cancer. Moreover, differentiated plasma cells were shown to produce antibodies against tumor antigens, which contributed to the immune protection in lung cancer patients [20]. As an example of cell-mediated immunity, Th17 cells produced interleukin-22, which promoted the growth, migration, and invasion of tumor cells [21, 22]. Interleukin-22 also induced tumor cells to express CD155, which inhibits the function of NK cells and supports tumor cell metastasis [23]. In this study, we found that SLC7A8 expression was associated with B cells, interstitial DCs, mast cells, CD56 bright cells, NK cells, plasmacytoid DCs, T follicular helper cells, Th2 cells, Th17 cells. Moreover, it was correlated with several immune factors, including CD5, CD34, CD36, CD37, CD79B, CDA, CDC7, CCL4, and CCL28. These results suggest that immune factors were significant in deciding the prognostic value of SLC7A8 in LUAD. We believe that elucidating the relationship between SLC7A8 and the immune microenvironment can lead to a breakthrough in the future treatment of LUAD.
Cancer is a disease characterized by dysregulated cell proliferation [24]. The knockdown of CYP2S1 effectively inhibited the migration, invasion, and proliferation of LUAD cells, while improving the OS of LUAD patients [25]. In this study, we found that the overexpression of SLC7A8 also effectively inhibited the proliferation and migration of LUAD cells. Bioinformatic analysis showed that SLC7A8 was closely associated with the cell cycle. Indeed, SLC7A8 inhibited the expression of the cell cycle-related proteins CDK1 and CCNB1. Therefore, SLC7A8 could check the progression of LUAD and improve its prognosis by influencing the proliferation and differentiation of LUAD cells.
This study has a couple of limitations. Firstly, although the relevant pathways were bioinformatically analyzed and experimentally verified, they were not validated through animal experiments, and we were unable to collect clinical samples for validation, which would have made the results more reliable. Secondly, we did not analyze extensive datasets, nor did we conduct multicenter cohort studies, Subsequent researchers may conduct multicenter studies based on this research.
This study demonstrated that the expression of SLC7A8 was attenuated in LUAD tissues. SLC7A8 can serve as a good prognostic marker for LUAD, and its overexpression inhibited the migration, invasion, and proliferation of LUAD cells.
Materials and Methods
Data of LUAD patients
SLC7A8 expression data in LUAD tissues and normal lung tissues were downloaded from TCGA and Xena. TCGA included 594 samples and Xena included 862 samples from the Genotype-Tissue Expression and TCGA databases. The clinical pathological and prognostic survival data of 522 LUAD patients were also downloaded from TCGA.
Identifying SLC7A8 expression in LUAD
The expression levels of SLC7A8 in normal lung tissues and LUAD tissues were determined by performing expression analysis on TPM data from TCGA and Xena. The expression of SLC7A8 in human normal lung epithelial cell and LUAD were determined by Western blotting.
Evaluating the clinical value of SLC7A8 in LUAD
After the LUAD patients were divided into groups according to their clinical pathological features, the expression levels of SLC7A8 in LUAD tissues were determined based on TPM data from TCGA. The clinical diagnostic value of SLC7A8 in LUAD was analyzed using ROC curves. Kaplan–Meier analysis and Cox regression were used to assess the prognostic value of SLC7A8 in LUAD, and prognostic nomograms were constructed based on it.
Cell culture
The human lung epithelial cell line BEAS-2b was purchased from Abclonal (Abclonal, China). The LUAD cell lines A549 and H1299 were purchased from American Type Culture Collection (ATCC, China). BEAS-2b cells were cultured in DMEM (Gibco, Thermo Fisher Scientific, Inc., USA) with 10% FBS (Gibco, Thermo Fisher Scientific, USA) and 1% Penicillin-Streptomycin solution at 37°C with 5% CO2. The LUAD cell lines were cultivated in Roswell Park Memorial Institute-1640 medium at 37°C with 5% CO2.
Plasmid transfection
The SLC7A8 expression plasmid and control vector were synthesized by RiboBio (China). A549 and H1299 cells were cultured in 6-well plates to about 80% confluence and transfected with 2 μg of plasmid using Lipofectamine®3000 (Thermo Fisher Scientific, USA) following standard protocols. After 6–8 h, the culture medium was replaced with a standard medium.
Western blotting
Total protein was extracted from A549, H1299, and BEAS-2b cells using RIPA buffer (Servicebio, China) and quantified using a BCA kit (Servicebio, China). The proteins were resolved by SDS–PAGE (Servicebio, China), transferred to PVDF membranes (Millipore, USA), blocked with 5% milk powder, probed overnight at 4°C with anti-SLC7A8 (1:1000, Abclonal, China), anti-CCNB1 (1:1000, Abclonal, China), anti-CDK1 (1:1000, Abclonal, China), and anti-β-actin (1:1000, Proteintech, China), incubated at room temperature for 1 h with anti-Rabbit IgG (1:3000, Jackson, USA) and anti-Mouse IgG (1:3000, Jackson, USA) secondary antibodies, washed with Tris-buffered saline with Tween 20, and finally detected using an ECL kit (Beyotime, China).
Cell proliferation
The proliferation of A549 and H1299 cells was assessed using the CCK-8 and colony formation assays [26, 27]. For the CCK-8 assay, the transfected cells were seeded in 96-well plates and incubated with 10 μL of CCK-8 solution for 2 h. The absorbance was measured at 450 nm using a microplate reader at 24, 48, 72, and 96 h. For the colony formation assay, the transfected cells were seeded in 6-well plates, incubated for 14 days, and fixed for 1 h. The colonies were then photographed and counted.
Wound healing assay
Cells were seeded in 6-well plates and cultured for 12 h to reach 100% confluence. The “wound” was made by scratching the monolayer with a sterile 200-μL pipette tip. Photos were taken immediately (0 h) and 24 h later at 100× magnification. The healing rate (healing width at 0 h/healing width at 24 h) was calculated using ImageJ.
Transwell assay
The migration and invasion of A549 and H1299 cells were assessed using the Transwell assay [28]. To assess cell migration, the cells were cultured to the logarithmic growth phase, washed once each with phosphate-buffered saline and serum-free medium, suspended in serum-free medium at a concentration of 1 × 105/mL, and seeded in the upper chamber of a 24-well Transwell chamber (Biosciences, USA). The lower chamber contained 600 μL of Roswell Park Memorial Institute-1640 medium containing 20% fetal bovine serum. After incubating for 24 h at 37°C, the lower chamber was fixed in 70% methanol for 1 h and stained with crystal violet. The cells in the low chamber were photographed and calculated. The protocol of the invasion assay was the same as that of the migration assay, except for the application of 100 μL of Matrigel to the upper surface of the membrane.
Correlation between SLC7A8 expression and immune infiltration in LUAD
LUAD tissues from TCGA were evaluated using single-sample GSEA, and the relationships between SLC7A8 expression and immune cell infiltration were analyzed using Spearman’s correlation. Moreover, this method was applied to explore the relationships between SLC7A8 expression and immune factors.
Statistical analysis
SLC7A8 expression in LUAD tissues was investigated using the t-test [29]. The diagnostic value of SLC7A8 in LUAD was analyzed by computing the area under the ROC curves. The prognostic value of SLC7A8 was ascertained using Cox regression and Kaplan–Meier survival analysis. The survival curves were constructed using the median grouping method, where 50% of patients were defined as having high expression of SLC7A8 [30]. Correlation analysis was used to investigate the relationship between SLC7A8 expression and immune cell infiltration in LUAD. P < 0.05 was considered to be significant.
Availability of data and materials
The datasets used during the current study are available from the corresponding authors upon reasonable request.
Author Contributions
Conception and design were done by Jiu-Ling Chen and Feng Zhao. Data acquisition and analysis was done by Fang-Ming Wang. Fang-Ming Wang, Li-Qiang Xu, and Zhong-Chao Zhang drafted the manuscript. Jiu-Ling Chen, Li-Qiang Xu, Qiang Guo, Zhi-Peng Du, Yue Lei, Chuang-Yan Wu, and Feng Zhao, revised the manuscript. The experiments were done by Zhi-Peng Du and Zhong-Chao Zhang, who contributed equally to this work. All authors have read and agreed to the final version of the manuscript.
Acknowledgments
We acknowledge the participants and clinicians who contributed data to this study.
Conflicts of Interest
The authors declare no conflicts of interest related to this study.
Ethical Statement
All clinical information and data were obtained from public databases or purchased from commercial sources, and therefore, the ethical approval is not required for this study. All procedures performed in this study were in accordance with relevant research regulations.
Funding
This work was supported by the National Natural Science Foundation of China (No. 82100299).
References
- 1. Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, Bray F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin. 2021; 71:209–49. https://doi.org/10.3322/caac.21660 [PubMed]
- 2. Chen Z, Fillmore CM, Hammerman PS, Kim CF, Wong KK. Non-small-cell lung cancers: a heterogeneous set of diseases. Nat Rev Cancer. 2014; 14:535–46. https://doi.org/10.1038/nrc3775 [PubMed]
- 3. Zhang L, Zhang Y, Wang C, Yang Y, Ni Y, Wang Z, Song T, Yao M, Liu Z, Chao N, Yang Y, Shao J, Li Z, et al. Integrated single-cell RNA sequencing analysis reveals distinct cellular and transcriptional modules associated with survival in lung cancer. Signal Transduct Target Ther. 2022; 7:9. https://doi.org/10.1038/s41392-021-00824-9 [PubMed]
- 4. He L, Chen J, Xu F, Li J, Li J. Prognostic Implication of a Metabolism-Associated Gene Signature in Lung Adenocarcinoma. Mol Ther Oncolytics. 2020; 19:265–77. https://doi.org/10.1016/j.omto.2020.09.011 [PubMed]
- 5. Breitenecker K, Homolya M, Luca AC, Lang V, Trenk C, Petroczi G, Mohrherr J, Horvath J, Moritsch S, Haas L, Kurnaeva M, Eferl R, Stoiber D, et al. Down-regulation of A20 promotes immune escape of lung adenocarcinomas. Sci Transl Med. 2021; 13:eabc3911. https://doi.org/10.1126/scitranslmed.abc3911 [PubMed]
- 6. El Ansari R, Alfarsi L, Craze ML, Masisi BK, Ellis IO, Rakha EA, Green AR. The solute carrier SLC7A8 is a marker of favourable prognosis in ER-positive low proliferative invasive breast cancer. Breast Cancer Res Treat. 2020; 181:1–12. https://doi.org/10.1007/s10549-020-05586-6 [PubMed]
- 7. Luo X, Yin P, Reierstad S, Ishikawa H, Lin Z, Pavone ME, Zhao H, Marsh EE, Bulun SE. Progesterone and mifepristone regulate L-type amino acid transporter 2 and 4F2 heavy chain expression in uterine leiomyoma cells. J Clin Endocrinol Metab. 2009; 94:4533–9. https://doi.org/10.1210/jc.2009-1286 [PubMed]
- 8. Min Q, Wang Y, Wu Q, Li X, Teng H, Fan J, Cao Y, Fan P, Zhan Q. Genomic and epigenomic evolution of acquired resistance to combination therapy in esophageal squamous cell carcinoma. JCI Insight. 2021; 6:e150203. https://doi.org/10.1172/jci.insight.150203 [PubMed]
- 9. Asada K, Kobayashi K, Joutard S, Tubaki M, Takahashi S, Takasawa K, Komatsu M, Kaneko S, Sese J, Hamamoto R. Uncovering Prognosis-Related Genes and Pathways by Multi-Omics Analysis in Lung Cancer. Biomolecules. 2020; 10:524. https://doi.org/10.3390/biom10040524 [PubMed]
- 10. Siegel RL, Miller KD, Fuchs HE, Jemal A. Cancer Statistics, 2021. CA Cancer J Clin. 2021; 71:7–33. https://doi.org/10.3322/caac.21654 [PubMed]
- 11. Nakanishi K, Matsuo H, Kanai Y, Endou H, Hiroi S, Tominaga S, Mukai M, Ikeda E, Ozeki Y, Aida S, Kawai T. LAT1 expression in normal lung and in atypical adenomatous hyperplasia and adenocarcinoma of the lung. Virchows Arch. 2006; 448:142–50. https://doi.org/10.1007/s00428-005-0063-7 [PubMed]
- 12. Jiang S, Zou J, Dong J, Shi H, Chen J, Li Y, Duan X, Li W. Lower SLC7A2 expression is associated with enhanced multidrug resistance, less immune infiltrates and worse prognosis of NSCLC. Cell Commun Signal. 2023; 21:9. https://doi.org/10.1186/s12964-022-01023-x [PubMed]
- 13. Jahani M, Azadbakht M, Norooznezhad F, Mansouri K. l-arginine alters the effect of 5-fluorouracil on breast cancer cells in favor of apoptosis. Biomed Pharmacother. 2017; 88:114–23. https://doi.org/10.1016/j.biopha.2017.01.047 [PubMed]
- 14. Baba H, Kanda M, Sawaki K, Nakamura S, Ueda S, Shimizu D, Koike M, Kodera Y, Fujii T. SLC7A9 as a Potential Biomarker for Lymph Node Metastasis of Esophageal Squamous Cell Carcinoma. Ann Surg Oncol. 2022; 29:2699–709. https://doi.org/10.1245/s10434-021-11001-1 [PubMed]
- 15. Tang B, Xu W, Wang Y, Zhu J, Wang H, Tu J, Weng Q, Kong C, Yang Y, Qiu R, Zhao Z, Xu M, Ji J. Identification of critical ferroptosis regulators in lung adenocarcinoma that RRM2 facilitates tumor immune infiltration by inhibiting ferroptotic death. Clin Immunol. 2021; 232:108872. https://doi.org/10.1016/j.clim.2021.108872 [PubMed]
- 16. Liu L, Xu S, Huang L, He J, Liu G, Ma S, Weng Y, Huang S. Systemic immune microenvironment and regulatory network analysis in patients with lung adenocarcinoma. Transl Cancer Res. 2021; 10:2859–72. https://doi.org/10.21037/tcr-20-2275 [PubMed]
- 17. Shen Y, Hou N, Han F, Chen B, Shi J, Sun X. Comprehensive Analysis of Tumor Immune Microenvironment and Prognosis of m6A-Related IncRNAs in Lung Adenocarcinoma. Crit Rev Eukaryot Gene Expr. 2022; 32:77–91. https://doi.org/10.1615/CritRevEukaryotGeneExpr.2022042417 [PubMed]
- 18. Kiriu T, Yamamoto M, Nagano T, Hazama D, Sekiya R, Katsurada M, Tamura D, Tachihara M, Kobayashi K, Nishimura Y. The time-series behavior of neutrophil-to-lymphocyte ratio is useful as a predictive marker in non-small cell lung cancer. PLoS One. 2018; 13:e0193018. https://doi.org/10.1371/journal.pone.0193018 [PubMed]
- 19. He J, Ding H, Li H, Pan Z, Chen Q. Intra-Tumoral Expression of SLC7A11 Is Associated with Immune Microenvironment, Drug Resistance, and Prognosis in Cancers: A Pan-Cancer Analysis. Front Genet. 2021; 12:770857. https://doi.org/10.3389/fgene.2021.770857 [PubMed]
- 20. Germain C, Gnjatic S, Tamzalit F, Knockaert S, Remark R, Goc J, Lepelley A, Becht E, Katsahian S, Bizouard G, Validire P, Damotte D, Alifano M, et al. Presence of B cells in tertiary lymphoid structures is associated with a protective immunity in patients with lung cancer. Am J Respir Crit Care Med. 2014; 189:832–44. https://doi.org/10.1164/rccm.201309-1611OC [PubMed]
- 21. Kryczek I, Lin Y, Nagarsheth N, Peng D, Zhao L, Zhao E, Vatan L, Szeliga W, Dou Y, Owens S, Zgodzinski W, Majewski M, Wallner G, et al. IL-22(+)CD4(+) T cells promote colorectal cancer stemness via STAT3 transcription factor activation and induction of the methyltransferase DOT1L. Immunity. 2014; 40:772–84. https://doi.org/10.1016/j.immuni.2014.03.010 [PubMed]
- 22. Kobold S, Völk S, Clauditz T, Küpper NJ, Minner S, Tufman A, Düwell P, Lindner M, Koch I, Heidegger S, Rothenfuer S, Schnurr M, Huber RM, et al. Interleukin-22 is frequently expressed in small- and large-cell lung cancer and promotes growth in chemotherapy-resistant cancer cells. J Thorac Oncol. 2013; 8:1032–42. https://doi.org/10.1097/JTO.0b013e31829923c8 [PubMed]
- 23. Briukhovetska D, Suarez-Gosalvez J, Voigt C, Markota A, Giannou AD, Schübel M, Jobst J, Zhang T, Dörr J, Märkl F, Majed L, Müller PJ, May P, et al. T cell-derived interleukin-22 drives the expression of CD155 by cancer cells to suppress NK cell function and promote metastasis. Immunity. 2023; 56:143–61.e11. https://doi.org/10.1016/j.immuni.2022.12.010 [PubMed]
- 24. Evan GI, Vousden KH. Proliferation, cell cycle and apoptosis in cancer. Nature. 2001; 411:342–8. https://doi.org/10.1038/35077213 [PubMed]
- 25. Guo H, Zeng B, Wang L, Ge C, Zuo X, Li Y, Ding W, Deng L, Zhang J, Qian X, Song X, Zhang P. Knockdown CYP2S1 inhibits lung cancer cells proliferation and migration. Cancer Biomark. 2021; 32:531–9. https://doi.org/10.3233/CBM-210189 [PubMed]
- 26. Liu Y, Liang L, Ji L, Zhang F, Chen D, Duan S, Shen H, Liang Y, Chen Y. Potentiated lung adenocarcinoma (LUAD) cell growth, migration and invasion by lncRNA DARS-AS1 via miR-188-5p/ KLF12 axis. Aging (Albany NY). 2021; 13:23376–92. https://doi.org/10.18632/aging.203632 [PubMed]
- 27. Huo X, Wang H, Huo B, Wang L, Yang K, Wang J, Wang L, Wang H. FTX contributes to cell proliferation and migration in lung adenocarcinoma via targeting miR-335-5p/NUCB2 axis. Cancer Cell Int. 2020; 20:89. https://doi.org/10.1186/s12935-020-1130-5 [PubMed]
- 28. Liang J, Li H, Han J, Jiang J, Wang J, Li Y, Feng Z, Zhao R, Sun Z, Lv B, Tian H. Mex3a interacts with LAMA2 to promote lung adenocarcinoma metastasis via PI3K/AKT pathway. Cell Death Dis. 2020; 11:614. https://doi.org/10.1038/s41419-020-02858-3 [PubMed]
- 29. Guo Q, Wu CY, Jiang N, Tong S, Wan JH, Xiao XY, Mei PY, Liu HS, Wang SH. Downregulation of T-cell cytotoxic marker IL18R1 promotes cancer proliferation and migration and is associated with dismal prognosis and immunity in lung squamous cell carcinoma. Front Immunol. 2022; 13:986447. https://doi.org/10.3389/fimmu.2022.986447 [PubMed]
- 30. Yousef GM, Scorilas A, Katsaros D, Fracchioli S, Iskander L, Borgono C, Rigault de la Longrais IA, Puopolo M, Massobrio M, Diamandis EP. Prognostic value of the human kallikrein gene 15 expression in ovarian cancer. J Clin Oncol. 2003; 21:3119–26. https://doi.org/10.1200/JCO.2003.09.111 [PubMed]