



Introduction
Sepsis is a potentially fatal organ failure brought on by an improperly controlled host response to infection [1], with a high morbidity and mortality rate worldwide. In 2017, the World Health Assembly listed sepsis as a global health priority [2]. Sepsis is one of the leading causes of death in the intensive care unit (ICU) [3]. According to statistics, sepsis accounts for 20% of annual deaths worldwide [4, 5]. Sepsis has a very complicated etiology that involves pathophysiological processes such as an excessive inflammatory response, pyroptosis, immunological dysfunction, mitochondrial damage, coagulation failure, oxidative stress, apoptosis, and autophagy, ultimately leading to organ dysfunction [6–10]. Over the past few decades, there have been efforts to come up with sepsis treatment strategies. In recent years, much progress has been achieved in the anti-infective, fluid resuscitation, hemodynamic sustain and organ function support therapy of sepsis using microbiology facilities and nanotechnology drug delivery platforms [11, 12]. However, the current clinical management of septic patients is still supportive rather than curative. It must be acknowledged that sepsis is a challenging issue for ICU physicians to overcome due to its multi-causal nature. Therefore, it is essential to study the molecular mechanisms underlying the biological processes involved in sepsis in order to optimize treatment options for the condition.
To date, there are more than 100 recognized modifications involved in regulating the bio-metabolic processes of RNA [13]. The most well studied RNA modification to date is N6-methyladenosine (m6A) methylation. Human messenger RNAs (mRNA), ribosomal RNAs (rRNA), and small nuclear RNAs (snRNA) all carry the m6A modification. m6A methylation is a reversible posttranscriptional modification of mRNA and regulates mRNA biogenesis and function [14]. Such modification regulates multiple steps of RNA processing including splicing, export, localization, decay and translation. More than 12,000 m6A loci were found in more than 7,000 human gene transcripts using antibody-mediated capture and massively parallel sequencing-based m6A-seq [15]. Studies have shown that m6A modification-related proteins are strongly associated with disease severity and prognosis [16, 17]. Especially, m6A methylation plays an essential role in inflammation by regulating three inflammatory signaling pathways including MAPK, JAK/STAT3, and PI3K [18]. Further data have demonstrated that METTL14-mediated m6A methylation negatively regulates inflammatory response in the context of sepsis [19]. It was discovered that insulin-like growth factor 2 mRNA binding proteins (IGFBPs), one of the m6A methylation binding proteins, influence the initial stages of septic shock [20]. Moreover, entire or RNA-specific therapeutic treatment of m6A methylation dynamics may be helpful to prevent and mitigate sepsis-induced disseminated intravascular coagulation [21]. In general, growing data suggests that m6A methylation is a mechanism that affects the onset and progression of sepsis. However, the regulatory function and underlying mechanisms of m6A in sepsis haven’t been thoroughly illuminated.
Under physiological environments, autophagic activity is often limited and serves as the cell’s guardian. However, when cells are exposed to outside stimuli such as pathogenic bacteria, hypoxia, and endotoxins, autophagic activity is significantly increased. Clearly, defects in the selective regulation of autophagy may lead to disease [22]. Autophagy is activated in the early stages of sepsis, followed by a phase of impaired autophagy [23]. Previous researches have shown that autophagy activation during sepsis is crucial for preventing subsequent lung, renal, and cardiac injury [24–26]. Existing research recognizes the critical role played by m6A methylation modification-related proteins in the biogenesis of autophagy [27, 28]. There is a growing awareness about the biological significance of the m6A modification on the transcription and translation of genes related to autophagy as well as the overall impact of conferring RNA specificity [29].
Clinically, m6A methylation-modified autophagy mechanisms play a crucial role in improving the prognosis of patients with sepsis [30]. There is no doubt that the connection between m6A methylation and autophagy will offer fresh perspectives on the management of sepsis. However, the effect of m6A-modified autophagy in the pathophysiology of sepsis remains largely unclear. Therefore, it is essential to clarify the potential mechanisms that revealing the exact biological processes and specific organ function-protective roles of m6A-modified autophagy in sepsis. The relative evidence that supports whether m6A methylation modified autophagy influences the pathophysiological mechanisms of sepsis is compiled in this review. The graphical abstract of this study is shown in Figure 1.
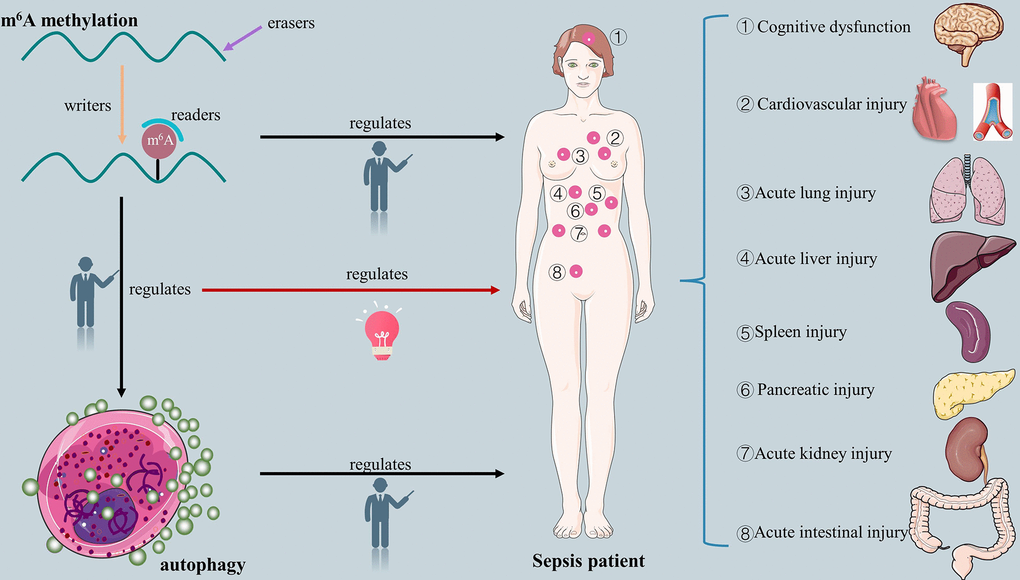
Figure 1. The overview of relationship between m6A methylation, autophagy, m6A-modified autophagy and sepsis.
m6A methylation
What is m6A methylation?
m6A methylation dynamically regulates RNA splicing, transport, localization, stability and translation [31]. m6A methylation, one of the common base modifications of mRNA, predominantly distributed in near stop codons, in 3’ UTRs [32] and within unusually long internal exons [15]. The blocked 5’terminal structure of heterogeneous nuclear RNAs, which exhibits striking similarities to one of the two varieties of blocked 5’ sequences seen in mRNAs, was discovered to be the site of m6A methylation as early as 1975 [33, 34]. Then, it has been discovered that the 5′ UTR’s m6A functions to stimulate mRNA translation when cells are under stress by taking the place of the 5’cap (which is the first step of most mRNAs translation) [35]. Of note, m6A is primarily found within the highly conserved consensus motif known as RRACH (R=G or A, H=A, C or U) in the majority of RNAs [36]. And then, m6A methylation also presences in a sequence context of UAC-(m6A)-GAGAA on top of a hairpin structure in transcript methionine adenosyltransferase 2A (MAT2A), which is mediated by methyltransferase-like 16 (METTL16) [37]. It is obvious that m6A methylation is an adenosine methylation at position N6, namely methylation of the sixth nitrogen atom on the RNA molecule adenosine. S-adenosylmethionine (SAM) provides nearly all the methyl groups necessary for cellular methylation reactions. The splicing of the MAT2A retained intron maintains high levels of intracellular SAM [38]. The specific mechanisms are as follows: the restriction of SAM prevents METTL16 from effectively inducing m6A methylation, which lengthens the time that it spends on a conserved hairpin (hp1) of MAT2A and promotes the splicing of retention intron, further provides enough SAM for m6A methylation.
Who was involved in the m6A methylation?
There are three crucial m6A methylation modification-related proteins exist in the process of m6A methylation: m6A methyltransferases (writers), m6A demethylases (erasers) and m6A recognition factors (readers) [39]. According to recent studies, writers include methyltransferase-like 3 (METTL3), methyltransferase-like 14 (METTL14), methyltransferase-like 5 (METTL5), METTL16, Wilms tumor 1-associated protein(WTAP), Vir-like m6A methyltransferase associated (VIRMA), RNA binding motif protein 15 (RBM15) and zinc-finger CCHC domain-containing protein 4 (ZCCHC4), erasers include fat mass and obesity-related proteins (FTO) and alkB homolog 5 (ALKBH5), readers include the YTH structural domain family (YTHDF) 1-3, YTH structural domain containing family (YTHDC) 1-2, eukaryotic translation initiation factor 3 subunit A(eIF3), insulin-like growth factor 2 mRNA binding protein 1/2/3(IGF2BP1/2/3), heterogeneous nuclear ribonucleoprotein A2/B1(HNRNPA2/B1) and HNRNPG, HNRNPC [40–42]. The potential role of m6A methylation modification-related proteins on RNA metabolism and the outcome of disease are summarized in Supplementary Table 1 [43–79].
How do m6A methylation modification-related proteins function during m6A methylation?
The METTL3-METTL14 compound is the ultimate important component in the writer proteins of m6A methylation. It is well established that the incidence of m6A methylation modification of various RNAs in mammals is inextricably linked to the activation of METTL3 and METTL14 [80]. In the process of m6A modification, METTL3 primarily functions as a catalytic core, while METTL14 provides a binding platform for RNA [81]. A recent study has shown that SUMOylation of METTL3 inhibits its m6A methyltransferase activity on RNAs [82]. The m6A methyltransferase’s core subunit, METTL14, works in stable heterodimer with METTL3 to catalyze m6A modification [83]. Subsequent research revealed that the methyltransferase activity of METTL3-METTL14 could only be elicited by the solution structure of the METTL3 zinc finger domain and not by the structural field of heterodimer between METTL14 and METTL3 [84]. Moreover, the METTL3-METTL14 complex is enlisted by WTAP, a regulatory subunit of the m6A methyltransferase complex, into nuclear patches that are enriched in pre-mRNA processing factors [85]. Prior works have also demonstrated that the mTORC1 modulates m6A methylation through regulating WTAP level to trigger the translational machinery for cell growth and proliferation [57, 86]. Contrary to the METTL3-METTL14 heterodimer, METTL16 is a single-component enzyme whose disordered loop is required to catalyze m6A methylation and whose N-terminal module is necessary for RNA binding [87]. Clearly, METTL16 in the cytoplasm and the nucleus serve different biological purposes. Only MAT2A mRNA and U6 snRNA were previously reported to directly deposit m6A from METTL16 [38, 88]. Reduced MAT2A mRNA degradation results from METTL16 localizing to hairpin 1 (hp1) on the 3’UTR of MAT2A mRNA and inducing MAT2A retained intron splicing [88]. In recent years, more RNAs with METTL16-mediated m6A methylation were reported in Supplementary Table 1 [43–79]. Additionally, wang, Fei et al. found that METTL16 facilitates mRNA 5 ‘cap-eIF4E recognition by sequestering eIF4E2 (translation initiation factor) [89]. Further studies suggest that this process of METTL16-mediated protein translation is independent of methyltransferase activity [89]. Therefore, in addition to catalyzing m6A methylation in the nucleus, METTL16 also participates in protein translation in the cytoplasm. The catalytic subunit m6A-METTL complex (MAC) and the regulatory subunit m6A-METTL-associated complex (MACOM) make up the m6A “writer”. The primary mechanism by which MACOM attaches to MAC is an interaction between WTAP and METTL3. Strikingly, WTAP and VIRMA comprise the basic structure of MACOM [90]. VIRMA recruits the METTL3/METTL14/WTAP, which are catalytic core components, to guide m6A methylation in 3’UTR and near stop codon of mRNA [91]. RBM15 plays a similar role to VIRMA in the methylation of m6A. RBM15 mediates m6A modification of targeted RNAs by targeting METTL3-METTL14 heterodimers to sites with or adjacent to m6A sites across the transcriptome [92]. ZCCHC4, a novel m6A methyltransferase that catalyzes m6A modification on rRNAs via binding to S-adenosyl-L-homocysteine, was recently reported [93]. ZCCHC4 is localized to the nucleolus, and ZCCHC4-mediated rRNA modification may also involve mRNA translation [94]. Further research established that ZCCHC4’s unique structural and enzymatic properties—namely, the formation of a complete RNA-binding surface by the association of the methyltransferase structural domain with the N-terminal GRF-type and C2H2 zinc finger structural domains and the C-terminal CCHC structural domain—are responsible for of its catalytic effect on rRNAs m6A modification [95].
m6A erasers predominantly catalyze demethylation of m6A-containing RNA. Two m6A demethylases have received a lot of attention to date: FTO and ALKBH5. FTO, one m6A eraser, primarily regulates the m6A modification in the nucleoplasm. FTO regulates pre-mRNA processing via its demethylation activity, which also influences mRNA stability close to the 7-methylguanosine cap, promotes cap-independent translation initiation at the 5’UTR, encourages exon jumping and alternative splicing at the pre-mRNA body, and modulates alternative poly(A) sites (APA) usage and 3’UTR length at the 3’UTR [96]. For instance, FTO can demethylate GAP-43 mRNA, and demethylation of GAP-43 mRNA may promote axonal elongation and regulate neural development [97]. ALKBH5, the other m6A eraser, localizes to nuclear speckles that are in charge of assembling mRNA processing factors. ALKBH5, a 2-oxoglutarate (2OG) and ferrous iron-dependent nucleic acid oxygenase (NAOX), has the potential to specifically bind single-stranded RNA attributed to a large loop (βIV–V) region that resembles the L1 loop of FTO [98]. ALKBH5’s demethylation activity has a momentous impact on gene expression, metabolism, and export of nuclear RNA (mainly mRNA), which regulates the biogenesis of m6A methylation on RNA [99]. In recent years, the crucial role of de-methylating of ALKBH5 in improving RNA stability has been particularly emphasized [100]. The demethylation of m6A modifications, exhibited by ALKBH5, improves the stability and expression levels of downstream RNAs that modulate heart regeneration and tumorigenicity [101, 102]. Does the m6A eraser’s demethylation activity work on methylated RNAs, though? A study offers an explanation: ALKBH5 and FTO keep their regulatory sites in an unmethylated stable state rather than reversing the methylated RNAs [103].
The variety of structural domains that m6A binding proteins possess allow for the division of these proteins into different families. Here, we’ll start out by introducing YTHDFs, which contain the YTH structural domain. YTH domain is known to directly bind the m6A base of methylated RNA [104]. YTHDF1 primarily recognizes the m6A methylation site of the downstream mRNA at the 3’UTR [66]. Mechanically, YTHDF1 recruits the transcripts of m6A-modified RNAs to facilitate their translation initiation [105], YTHDF2 induces the degradation of m6A-modified RNAs to decrease their stability [106], and YTHDF3 regulates the m6A methylation of downstream signal RNAs in synergy with YTHDF1 or YTHDF2 [107]. Contrary to these conventional beliefs, studies have proposed a novel unified model of m6A function, in which all m6A bits combine all three DF paralogs (YTHDF1, YTHDF2 and YTHDF3) in a fundamentally similar manner, influencing the degradation of mRNA through the interaction of these three key redundant DF proteins [108]. They did not, however, turn up any evidence that would support their hypothesis that these three DF proteins cooperating together have a role in promoting mRNA translation. Second, we’ll introduce the overview of YTHHDCs in the present paper, which share the same YTH structural domain. m6A methylation modification is a significant modality of regulation in mRNA splicing. The fate of the transcripts in terms of splicing dynamics and alternative splicing may be determined by early m6A sedimentation [109]. According to biochemical, structural, and transcriptome-wide PAR-CLIP (photoactivated ribonucleic acid enhanced cross-linking and immunoprecipitation) investigations, YTHDC1 is a nuclear RNA-binding protein that is responsible for recruiting mRNA splicing factors for pre-mRNA. The result from the current study has demonstrated that YTHDC1 promotes SRSF3 but antagonizes SRSF10 binding to RNAs at the m6A methylation site, further triggering the initiation of mRNA splicing [110]. It has been proposed that YTHDC2 may interact with translation and decay mechanisms in the context of particular binding to m6A in order to boost translation effectiveness and reduce the mRNA abundance of its targets [111]. Third, the identification of m6A by IGF2BPs depends on the K homology (KH) structural domain. Such m6A reading proteins selectively bind m6A-containing RNA using the KH structural domain and its flanking regions [104]. IGF2BPs have been reported to support the stability, storage and translation of their target mRNA transcripts by identifying their consensus GG-(m6A)-C sequences [78, 112]. The selectivity with which the remaining m6A reading proteins activate m6A causes them to be clustered together. Such m6A reading proteins bind m6A-containing transcripts through a m6A switch mechanism because the m6A modification weakens Watson-Crick base pairing of RNA and makes it easier for m6A reading proteins to recognize single-stranded RNA motifs; simultaneously, hnRNPA2B1 can also bind m6A-containing RNA with specificity by using the RRM structural domain and its flanking regions [104]. HNRNPG, a novel m6A methylation binding protein, binds purine-rich regions exposed by m6A modified RNA using its low-complexity region, regulating gene expression and selective splicing [42]. eIF3 binds directly to the 5’UTR m6A site of mRNA in the cytoplasm, which is sufficient to recruit the 43S complex and initiate translation in the absence of the cap-binding factor eIF4E [35, 113].
It is still controversial what function m6A-related proteins have in the pathogenic and physiological processes of the disease. For example, analysis reports of TCGA data indicate that high expression of METTL3 is associated with unfavorable prognosis in CRC patients [114]. Conversely, clinical research has demonstrated that METTL14 deletion is related to a poor prognosis in patients with CRC [52]. By modification of m6A, METTL3 can also assist in regulating the cardiac homeostasis and hypertrophy [115]. WTAP was also identified as an independent predictor of prognosis for patients with hepatocellular carcinoma [58]. According to one study, testicular Leydig cells experienced an increase in m6A methylation modification of RNA due to the inhibition of FTO, which led to apoptosis [116]. By examining the expression of m6A-related regulators and the probability of overall survival in HNSCC, Yu, Dan et al. discovered IGF2BP2 to be an independent prognostic factor in patients [77]. To sum up, various regulations of m6A methylation modification-related proteins play a critical role in the corresponding modified RNAs’ transcription by affecting their splicing, export, translation, and stability, which ultimately influences the development of these modified RNAs-mediated diseases. The diagrammatic sketch of these regulatory mechanisms is shown in Figure 2. It follows that m6A methylation is expected to develop as a therapeutic target for human diseases.
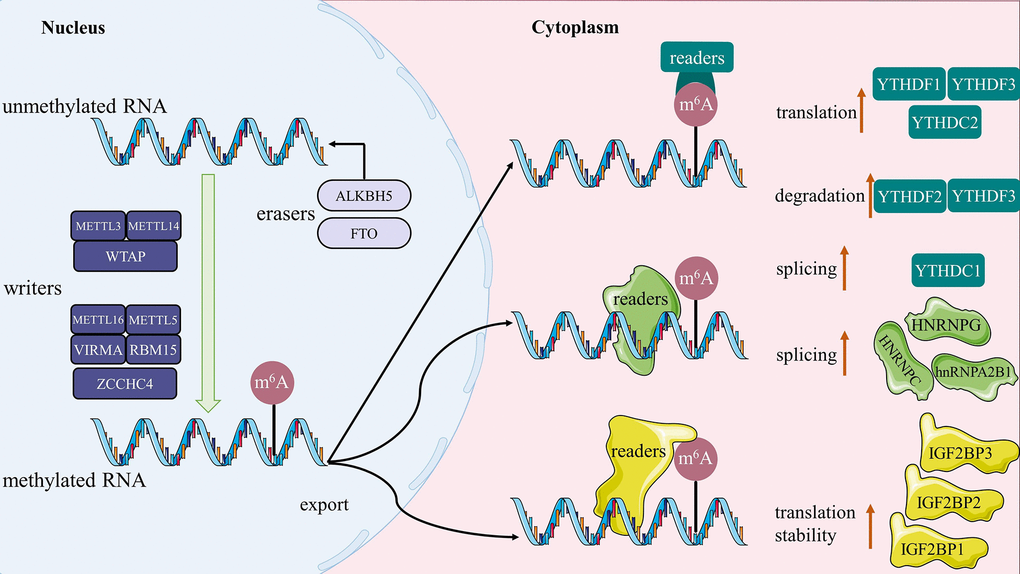
Figure 2. The sketch map of m6A methylation. Writers catalyze the m6A modification of RNA, erasers maintain the RNA in an unmethylated state, and the readers are ultimately responsible for determining the fate of the RNA (such as splicing, stability, degradation and translation).
However, as the field of study developed, we discovered that numerous m6A modification-related proteins interact to regulate the m6A methylation of targeted RNAs, rather than a single m6A modification-related protein, in the disease process. Many m6A methylation sites on SPRED2 mRNA have been found to be lost as a result of METTL3 deletion [117]. This impairs YTHDF1-mediated translation of the modified SPRED2 mRNA and increases NF-kB and STAT3 activation through the ERK pathway, which promotes tumor development and metastasis [117]. The level of SRY (sex determining region Y)-box 2 (SOX2) transcripts’ m6A methylation elevated as a result of METTL3. IGF2BP2 subsequently recognized methylated SOX2, maintaining its mRNA stability and expression. Ultimately, CRC development was triggered by high SOX2 expression [114]. However, when abundant in cellulose, METTL3’s role changes from catalyzing m6A methylation to promoting the initiation of mRNA translation. The production of dense polynucleotides, accelerated translation, and carcinogenic transformations all depend on METTL3-eIF3h interactions [118]. Moreover, another role m6A methylation frequently plays in disease development is targeted RNA degradation that is dependent on the METTL3-YTHDF2 interaction [73, 119]. METTL3 directs m6A modification of PKC-η, FAT4, and PDGFRA mRNAs to induce mRNA degradation via YTHDF2-dependent pathway, which promotes diabetes-related peripapillary cell dysfunction and stimulates retinal vascular complications [43]. As a result, METTL3 induces high levels of m6A modification in mRNA, and YTHDF2 identifies m6A sites in mRNA and promotes its degradation [120]. In the last resort, the synergistic effect of METTL3-YTHDF2 regulates the development of diseases via influencing disease-related genes expression. In a METTL3-FTO-dependent manner, m6A methylation plays the crucial role in the clinical and physiological processes of obesity cardiomyopathy, too [121]. ALKBH5 deletion leads to elevated m6A levels in downstream RNAs, and IGF2BP1 recognizes the exposed m6A sites and enhances their stability, thereby enhancing downstream RNA expression [122]. Similarly, ALKBH5-mediated m6A modification of its downstream targets is recognized by another m6A reader, YTHDF2, which is also responsible for degrading ALKBH5’s methylated downstream targets [123, 124]. Supplementary Table 1 [43–79] provides additional information on the essential role of writer/eraser-reader-dependent m6A methylation in the regulation of disease. In a word, m6A methylation is a dynamic and programmed process of RNAs modification. Even though each of the m6A modification-related proteins has a specific function, m6A modification of RNAs seems impossible to happen without the synergistic effect of these proteins. The level of RNAs’ m6A methylation in the nucleus is regulated by the m6A methyltransferases and demethylases, and the modified RNA enters the cytoplasm. Further affecting RNA splicing, degradation, stabilization, and translation are m6A binding proteins, which find and bind m6A residues on the transcript of the modified RNA. Therefore, this distinctive “writer/eraser-reader-dependent” paradigm for m6A methylation should be considered in the pathophysiological processes of disease, providing additional possibilities for therapeutic intervention.
Of course, the upstream signaling of m6A methylation-related proteins also influences their expression level, which in turn influences the level of downstream signaling that m6A regulates and, ultimately, the progression of the disease. For instance, Piwi-interacting RNA (piRNA)-14633 interacts with the 3’UTR of METTL14 to enhance the stability of METTL14 mRNA and encouraged the methylase activity of METTL14, promoting the m6A methylation levels of the downstream target (CYP1B1), and subsequently promoting the expression of CYP1B1, which in turn contributed to the oncogenesis of cervical carcinoma [53]. Additionally, by interacting with METTL3 and inhibiting its RNA methylation activity, cardiac-hypertrophy-associated piRNA (CHAPIR) prevents the m6A modification of PARP10 mRNA. This causes a blockage of the YTHDF2-mediated degradation of the PARP10 mRNA transcripts and an increase in PARP10 expression, which leads to cardiac hypertrophy [119].
The regulatory role of m6A methylation in sepsis
A complicated systemic inflammatory response, immunological dysfunction, aberrant coagulation, oxidative stress, apoptosis, dysregulation of autophagy, and tissue damage are all factors in the pathogenesis of sepsis. m6A methylation facilitates these biological processes by differentially regulating specific RNAs. Trials have demonstrated a substantial correlation between m6A regulators including ALKBH5, HNRNPC, KIAA1429, WTAP, and YTHDF2 and 28-day cumulative mortality in sepsis patients. Of note, HNRNPC, KIAA1429, and YTHDF2 are protective genes with a hazard ratio (HR) < 1, but ALKBH5 and WTAP are dangerous genes with a HR > 1 [30]. Further research also confirmed the protective effects of FTO, HNRNPC, YTHDC1, and RBM15B in sepsis patients [125]. In one animal study, increased expression of METTL3 and low expression of METTL14, ALKBH5, FTO, and YTHDF2 were found following lipopolysaccharide (LPS) induction. Subsequent research demonstrated that m6A modification plays a role in the pathophysiology of sepsis and mediates sepsis-induced liver injury [126]. The m6A modification may have an intimate and intricate interaction relationship with the cardiovascular injury generated by the different physio-pathological conditions of sepsis. Shen et al. observed that downregulation of METTL3 and WTAP was partially responsible for the decrease in major m6A levels in aortic RNA during sepsis [21]. Additionally, there was a significant decrease in the levels of m6A modification in septic cardiac tissue, indicating a critical role for m6A modification in the pathogenesis of sepsis-related myocardial damage [127]. In summary, the evidence that is now available generally points to the possibility that therapeutic adjustments of cellular m6A methylation may assist with alleviating secondary organ dysfunction during sepsis.
We can’t only look at the overall degree of change in m6A methylation in sepsis, though, given the complicated pathophysiological mechanisms of sepsis and the dynamic and multifactorial role of m6A methylation. As a result, it is appropriate to incorporate the “reader/eraser-reader” model of m6A modification into the mechanistic investigation of sepsis and to clarify the specific molecular mechanisms of sepsis in the context of the dynamic process of cellular m6A modification. Data mining revealed that the majority of m6A-RNA methylation regulators’ expression was down-regulated in sepsis, with only a few up-regulated [128]. Recently, several in vitro experiments with the sepsis model observed abundant m6A methylation in LPS-induced cardiomyocytes (H9C2). Mechanically, METTL3 catalyzed m6A modification of HDAC4 mRNA, and IGF2BP1 identified the m6A site on HDAC4 mRNA and strengthened its stability, which consequently stimulates the inflammatory damage of cardiomyocytes induced by sepsis [129]. Obviously, METTL3-mediated m6A modifications on transcripts of numerous inflammatory signaling pathways are responsible for the excessive inflammatory responses and pyroptosis [130, 131]. More specifically, endotoxin invasion stimulates m6A methylation of intracellular inflammatory factors IL-6 and TNF-α transcripts in response to myocardial inflammation in sepsis [132, 133]. Likewise, YTHDF2 recognizes METTL3-mediated m6A modification of SLC7A11 mRNA and promotes the degradation of SLC7A11 mRNA, ultimately leading to ferroptosis in sepsis-induced myocardial injury [134]. A recent study reported that METTL3-induced m6A modification on ferroptosis was involved in the pathogenesis of sepsis-associated acute lung injury [135]. Conversely, there is a decreased m6A level in sepsis-induced acute respiratory distress syndrome (ARDS) in vivo and in vitro. Functionally, YTHDF1 recognized and stabilized METTL3-mediated m6A-modified tripartite motif-containing (Trim59) mRNA to protect the vascular endothelium against barrier dysfunction and inflammatory responses, which inhibits the evolution of ARDS during sepsis [136]. FoxO1/NF-κB is a recognized inflammatory signaling pathway that mediates the inflammatory response by promoting the generation of the inflammasome NLRP3. Previous studies have demonstrated that inhibition of FTO mediates m6A modification of FoxO1 mRNA and reduces its expression, thereby suppressing the inflammatory response in septic shock [137]. Moreover, inflammatory signaling pathway TLR4/NF-κB is negatively regulated by SOCS1 and Spi2a. It is understood that SOCS1 and Spi2a mRNA stability as well as translation are improved by METTL14-YTHDF1-dependent m6A methylation to prevent the progression of sepsis [19, 138]. Therefore, “writer/eraser-reader-dependent” m6A methylation may be a regulator of sepsis progression. It is concluded that the alterations in m6A modification during sepsis are closely associated with ferroptosis, pyroptosis, inflammatory and immune responses. Meanwhile, autophagy as a protective mechanism in sepsis and more m6A methylation regulating its biological role in the pathophysiological processes of sepsis need to be elucidated.
The regulatory role of m6A methylation in autophagy
The regulatory role of m6A methylation in the development of autophagy must be established in order to gather evidence for the association between m6A methylation-modified autophagy and sepsis, which indicates that this RNA modification contributes essentially to the biological processes of autophagy initiation, extension, and maturation. Basal autophagy is tightly regulated by transcriptional and epigenetic mechanisms to preserve intracellular homeostasis. The epigenetic regulation of m6A in the autophagic process of human diseases has gradually come to light in recent studies [139]. In reviewing the literatures, considerable evidences were found on the association between m6A methylation and autophagy. The regulation of m6A methylation on autophagy can be negative or positive, which may be related to the different functions of modified RNAs during autophagy and the specific effect of m6A modification-related proteins on targeted RNAs.
The direct regulatory role of m6A methylation in autophagy
The m6A modification directly regulates maturity of autophagy by affecting the activity of the autophagy-related proteins. One could argue that this regulation is negative. The stimulation of the ULK1 complex, which is comprised of ULK1 and the noncatalytic subunits FIP200 and ATG13, is the first step in the onset of autophagy. ALKBH5 maintained FIP200 at unmethylated steady-state levels, and YTHDF2 was unable to induce degradation of FIP200 in the cytoplasm because it failed to recognize m6A residues on the FIP200 transcript, resulting in increased FIP200 expression and activation of the autophagic pathway [140]. FTO-YTHDF2-dependent m6A methylation regulates the biological process of autophagy on ULK1 mRNA through the same mechanism as aforementioned [141]. Transcription factor EB (TFEB) is necessary for lysosomal biogenesis and autophagy [142, 143]. Increased m6A methylation of TFEB mRNA and decreased m6A expression level of TFEB mRNA are caused by upregulation of METTL3 and downregulation of ALKBH5 in ischemic heart disease, which together prevent the maturation of autophagy [144]. ATGs are a class of regulatory proteins that are essential for the formation of autophagosomes. Reduced FTO-mediated m6A modification on ATGs has been demonstrated to activate autophagy [145]. Moreover, when FTO is silenced, YTHDF2 binds to m6A methylation-enriched ATG5 and ATG7 transcripts, causing mRNA to decay and protein production to decline. This prevents the formation of autophagosomes [146]. Even though previous studies claimed that up-regulation of FTO would prevent autophagy from maturing [147]. The evidence presented thus far supports the idea that at the level of gene metabolism where m6A methylation regulates autophagy, the “writer” is primarily responsible for catalyzing m6A modification of RNA, while the “eraser” maintains the RNA in an unmethylated stable state, and it is the “reader” that ultimately determines the fate of the RNA. Furthermore, METTL3-mediated m6A modification reduces ATG7 expression by weakening the stability of ATG7 mRNA, the autophagic process is ultimately blocked [148]. Taken together, there are no studies on negative regulation factors of autophagy in this context, but the negativity of m6A methylation directly regulates autophagy is primarily reflected in the interaction between m6A modification-related proteins that can ultimately down-regulate the expression of autophagy-positive regulation factors.
However, such regulation may also be positive. YTHDF1 contributed to the translation of ATG2A and ATG14 by binding to the m6A site of methylated ATG2A and ATG14 mRNA, thus facilitating autophagy [149, 150]. Beclin1 contributes to the elongation of the autophagosome membrane. Several lines of evidence suggest that METTL14-YTHDF1-dependent m6A modification appears to trigger autophagy activation by stabilizing Beclin1 mRNA [151]. Consistently, METTL14-IGF2BPs-dependent m6A methylation plays the same role on Beclin1-mediated autophagy as above [152]. Moreover, down-regulated ALKBH5 promotes m6A methylation of Beclin1 and LC3 II/I mRNAs, resulting in the high expression of Beclin1 and LC3 II/I and activation of autophagy [153]. The presence of p62/SQSTM1-droplet, an autophagy selective receptor, creates a foundation for the formation of autophagosome [154], which may be related to recruiting more LC3. Therefore, the nuclear m6A methylation of SQSTM1 mRNA, which is mediated by YTHDC1, upregulates the expression of SQSTM1 and increases autophagic flux [76]. The evidences listed here suggest that the connection between proteins connected to m6A modification, which has the potential to up-regulate the expression of autophagy-positive regulation factors, is the main way that the positivity of m6A methylation directly regulates autophagy is manifested.
The indirect regulatory role of m6A methylation in autophagy
The m6A modification can also indirectly regulate maturity of autophagy by affecting the activity of the autophagy-related pathways. Additionally, there are both negative and positive correlations between m6A alterations and pathways involved in autophagy. We preferentially focus on expanding the conversation around negative regulation. Autophagy is positively regulated by the AMPK pathway in the progression of sepsis-induced cardiomyopathy [155]. Previous research has shown that m6A methylation inhibits autophagy by increasing the translation of protein phosphatase 1A (PPM1A), an AMPK negatively regulated factor, which is mediated by YTHDF1, while decreasing the stability and expression of calcium/calmodulin-dependent protein kinase kinase 2 (CAMKK2), an AMPK actively regulated factor, which is mediated by YTHDF2 [156]. They also found that a decrease in METTL3 and METTL14 and an increase in ALKBH5 in the process [156]. In the same way, our research goes further and focuses on the LKB1, an upstream kinase of AMPK. WTAP-mediated m6A methylation impaired the stability and expression of LKB1 mRNA, which prevents the AMPK pathway from being activated and inhibits autophagic flux [157]. Additionally, SIRT1 pathway activates autophagy by deacetylating a variety of ATGs. Mechanically, METTL14-dependent m6A modification mediates the degradation of SIRT1 mRNA, which provide a potential possibility for curbing autophagy [158]. On the other hand, the synergy of tumor protein p53 inducible nuclear protein 2 (TP53INP2) with LC3 and ATG7 is also critical for autophagy activation. FTO induces the decreased m6A modification of TP53INP2 transcript as well as high expression of TP53INP2, which ultimately promotes autophagy [159]. In response to extracellular stress, the FOXO3 pathway maintains cellular homeostasis by acting on high levels of intracellular ROS to mediate autophagy [160]. METTL3-induced m6A methylation decreases autophagic flux through enhancing FOXO3 RNA stability and expression in an YTHDF1-dependent manner [161]. Similar to the above, Rubicon is a cellular autophagy negative regulator gene that binds to Beclin1 to inhibit the autophagic pathway; METTL3-YTHDF1-dependent m6A methylation also decreases autophagic flux by enhancing the stability and translation of Rubicon mRNA [162]. Initial observations suggest that PI3K/AKT/mTOR signaling pathway plays an important role in the anti-autophagy effect [163]. IGF2BP3 promotes translation machinery associated 7 homolog (TMA7) mRNA stability and translation through recognition of the m6A site on the TMA7 3’-UTR, which in turn activates the PI3K/AKT/mTOR pathway and ultimately inhibits autophagy [164]. Conversely, activating transcription factor 4 (ATF4) negatively regulates the mTOR signaling pathway. High expression of FTO maintained ATF4 mRNA at unmethylated steady-state levels, and YTHDF2 was unable to induce degradation of ATF4 in the cytoplasm because it failed to recognize m6A residues on ATF4 mRNA transcripts, thus increasing ATF4 expression levels and activating the mTOR-suppressed autophagic pathway [165]. Overall, the m6A modification negatively regulates autophagic activity mainly by mediating the expression of upstream signaling pathways of autophagy regulators.
Then, we develop the discussion of positive regulation. ALKBH5-mediated reduction of m6A methylation indirectly inhibits the development of autophagy by regulating the expression of GSK3β/mTOR signaling pathway [166]. Earlier, study has observed that miR-199a impairs autophagy in thick heart muscle cells in a cell-autonomous way through targeted GSK3β/mTOR signal pathway [167]. More precisely, autophagy is negatively regulated by the GSK3β/mTOR signal pathway [168]. USP13 is known to be an essential deubiquitinase that stabilizes ATG5 by deubiquitination. Mechanically, USP13’s m6A modification is catalyzed by METTL3, and IGF2BP2 promotes USP13 mRNA translation by identifying m6A residues on USP13 transcripts, ultimately triggering autophagy [169]. Decapping Protein 2 (DCP2) is degraded as a result of METTL3’s induction of m6A methylation, which facilitates mitophagy via the Pink1-Parkin pathway [170].
To sum up, a sizable and intricate regulatory network of signaling pathways exists upstream of the autophagy regulators, and m6A modification in any member of this regulatory network will govern the bioprocess of autophagy by affecting the expression of its downstream signals. As shown in Figure 3, the aforementioned empirical findings in the present study provide a new understanding of m6A methylation regulates autophagy. First, we summarize the regulation of m6A methylation in the biological metabolism of autophagy as a “writer/eraser-reader-dependent” model, where the “writer” is mainly responsible for catalyzing the m6A modification of RNA, while the “eraser” maintains the RNA in an unmethylated state, and the “reader” is ultimately responsible for determining the fate of the RNA. Second, two approaches exist for m6A methylation to control autophagic activity: directly by altering the autophagic regulators themselves, or indirectly by altering the upstream signaling pathways that mediate the autophagic regulators’ expression. Then, m6A modification up-regulates autophagic activity by inhibiting the degradation of autophagy-negative factors or encouraging the expression of factors that promote autophagy. Conversely, it down-regulates autophagic activity. Last but not least, m6A modifications have an impact on autophagy regulators and autophagic activity by mediating the expression and degradation of the regulators’ upstream signaling pathways.
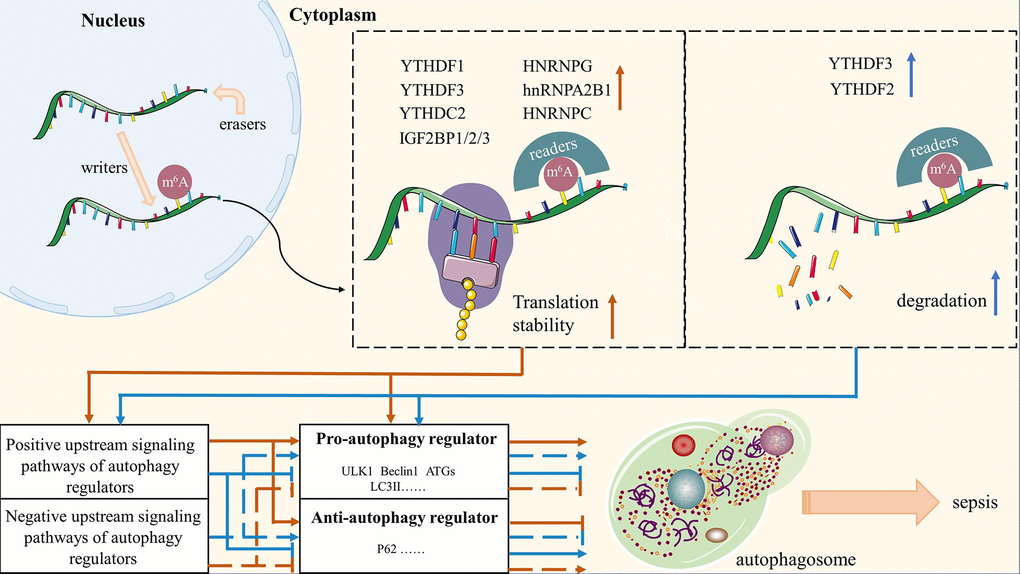
Figure 3. A summary of molecular mechanisms of m6A-modified autophagy on sepsis. Red arrows represent the role of m6A methylation in maintaining the stability of autophagy regulators and their upstream signaling pathways; Blue arrows represent the degradation of autophagy regulators and their upstream signaling pathways by m6A methylation.
The regulatory role of m6A methylation modified autophagy in sepsis
Autophagy is a conserved lysosomal degradation pathway that transports substrates (including large amounts of cytoplasm, organelles (e.g., mitochondria and peroxisomes), aggregation-prone proteins, and infectious agent) to lysosomes via double-membrane vesicles. The autophagy pathway plays homeostatic activities in protein and organelle quality control to maintain mammalian developmental and differentiation processes [171]. Although autophagy is generally regarded as an adaptable and protective biological process, it can be destroying when it occurs in excess or has defects. From the focus of studies on the biological functions of autophagy genes over the past two decades, autophagy specifically targets disease-causing proteins, intracellular microorganisms, and dysfunctional organelles; deficits in these processes that cause abnormal accumulations of inflammatory signals may be relevant to the pathophysiological mechanisms of inflammatory diseases [22]. Research shown that inhibiting autophagic flux increases the likelihood of non-canonical inflammasome pathways being activated, which impairs with the host’s ability to fight off infection [172]. Improved autophagic flux-based sepsis therapy options are suggested by a clinical investigation that reveals impaired autophagic flux in septic patients [173]. Our previous basic studies have revealed that activation of autophagy is protective of multiorgan function in a sepsis model [174, 175]. Then, cognitive dysfunction in sepsis-related encephalopathy is ameliorated by activation of PPAR-γ signaling pathway-mediated autophagy in astrocytes, as evidenced by high expression of LC3, ULK1, and low expression of P62 [176]. Conversely, inhibition of the autophagy pathway may likewise ameliorate sepsis-induced organic depression. In the pathogenesis of sepsis, for instance, it has become clear that suppressing autophagy mechanisms by targeting SIRT4, MAPKs, and Nrf2 pathways may be a useful strategy for protecting organ function [177–179]. Indeed, it has also been demonstrated that autophagy is activated early in the onset of sepsis, but that as the condition progresses, autophagic activity declines [180, 181]. There are opposing views in the existing literature on the crucial protective or destructive functions of autophagy in sepsis-induced organ damage. We speculate that the cause of this phenomenon might be connected to the respective alteration of post-transcriptional processes that autophagy regulators and their upstream signaling pathways go through during sepsis. Needless to say, the dynamic change of autophagy during the development of sepsis will continue to receive attention in the subsequent research. Consequently, the autophagy pathway appears to be closely involved in the pathogenesis of sepsis, and its modulation may be of therapeutic value in the clinical context. According to studies, the most common post-transcriptional modification is the m6A methylation, and the internal modifications it exerts in mRNAs are an intricate biological process [182]. Since the relationship between m6A methylation and autophagy has been elucidated in many human diseases [183], the molecular mechanisms by which m6A-autophagy interactions induce sepsis have received extensive attention from emergency physicians. The dual role of m6A modifications in sepsis is strikingly similar to that of autophagy, which can both promote and hinder the occurrence and development of sepsis [136, 184]. Furthermore, the mutual control of m6A modification and autophagy is becoming increasingly clear as a result of profound autophagy research, and their interactions can further affect the efficacy of sepsis therapy. Research on the sepsis-associated characteristic gene METTL3 has been increasingly prevalent in recent years, with the goal of exploring potential epigenetic treatment targets for sepsis patients. Under disease-related circumstances, the expression of METTL3 target genes can fluctuate, which affects the pathophysiology of inflammatory diseases by affecting the expression of downstream target genes [185]. A key pathogenic mechanism in sepsis, PINK1/Parkin-mediated mitochondrial autophagy, is negatively regulated by DCP2 [26]. The latest report indicates that the m6A methyltransferase METTL3 can facilitate mitochondrial autophagy mediated by the PINK1/Parkin pathway by triggering the m6A methylation of DCP2, which results in the degradation of DCP2 [170]. Additionally, it was discovered that METTL3-IGF2BP2-dependent m6A modification emerged as a contributing factor in the deterioration of sepsis-induced acute lung damage [184]. Likewise, such m6A modification means promote autophagy in certain diseases [186]. Further researches that METTL3-mediated m6A methylation inhibits the activation of autophagy also supported the anti-inflammatory function of this modulation in infectious illnesses [187]. More precisely, m6A modification leads to impaired autophagic flux ending in reduced cellular viability during sepsis-induced organ dysfunction [188]. Consequently, m6A methylation affects the regulation of autophagy during a dysregulated host response to infection in addition to being involved in the pathophysiology of sepsis and the development of autophagy. To summarize, m6A methylation modified autophagy may be the potential molecular mechanism and have clinical value in sepsis, but more research is needed.
Conclusions
In this review, we present a hypothesis on “writer/eraser-reader-dependent” m6A methylation-modified autophagy that may aid in the discovery of novel therapeutic targets to reduce morbidity and mortality related to organ dysfunction subsequent to sepsis and give a theoretical basis for more comprehensive management of sepsis patients. Globally, the health of people is seriously threatened by the complex series of diseases known as sepsis. Over the last few decades, we have been working hard to uncover the underlying molecular mechanisms in sepsis. As sepsis progresses, there is growing evidence that alterations in gene expression and epigenetic regulation are related to organ dysfunction. It is well known that one of the recognized RNA modifications that controls epigenetic and gene expression is m6A methylation. In addition, a series of investigations have demonstrated that m6A methylation regulates a variety of biological processes in sepsis. On the other hand, m6A methylation plays a crucial role in the post-transcriptional steps of genes, affecting the stability, export, splicing and translation of the transcripts involved in the autophagic process. Previous studies have also confirmed the importance of maintaining the orderly and complete autophagic process in the prevention of sepsis. Unfortunately, there is only limited evidence elucidating a potential relationship between m6A-modified autophagy and sepsis. For example, a recent study revealed that METTL3 mediates the m6A methylation of SIRT1 mRNA, which suppresses SIRT1 protein expression and autophagic flux and eventually results in sepsis-induced acute lung injury [188]. In conclusion, the novel regulatory model of m6A methylation modification proposed in this paper provides an innovative research direction for the therapies of sepsis.
The following are some of the key points that needs to be covered in this article: (1) A gene’s whole post-transcriptional biological process is impacted by m6A methylation, and each of the proteins involved in this modification has a specific function in this process. By summarizing the current evidence on m6A modification-related proteins acting on the metabolism of RNA, we emphasize the importance of synergistic interactions among m6A modification-related proteins in regulating m6A methylation during the pathophysiology of disease. (2) We suggest a novel m6A modification model-“writer/eraser-reader-dependent” m6A methylation (Figure 2), and thoroughly analyze the specific molecular mechanisms by which it regulates sepsis. (3) At the level of gene metabolism where m6A methylation regulates autophagy, the “writer” is mainly responsible for catalyzing the m6A modification of RNA, while the “eraser” maintains the RNA in an unmethylated state, and the “reader” is ultimately responsible for determining the fate of the RNA. (4) In order provide new insights into the regulatory role of m6A modifications throughout the autophagic process, we have meticulously collated relevant potential mechanisms between m6A modifications and autophagy, including direct evidence for particular molecular mechanisms and indirect evidence for pertinent signaling pathways (Figure 3). (5) We speculate that the pathophysiology of sepsis may also be influenced by the putative molecular mechanisms between m6A modification and autophagy. However, there is still no clear evidence for the effect of m6A-modified autophagy on sepsis, and further exploration of potential links between the listed mechanisms is required.
Supplementary Materials
Author Contributions
LS Y and JF Z performed study concept and design; CF B performed drawing of mechanism diagrams and writing, review and revision of the paper; JL and XD H provided acquisition, analysis and interpretation of materials. All authors have reviewed the final version of the manuscript and approved it for publication.
Acknowledgments
The authors thank Department of Emergency Medical, General Hospital of Ningxia Medical University for supporting this work.
Conflicts of Interest
All authors declare that there are no conflicts of interest.
Funding
This review is supported by research grants from the National Natural Science Foundation of China (82205116) and the Natural Science Fund of Ningxia (2022AAC03593).
References
- 1. Singer M, Deutschman CS, Seymour CW, Shankar-Hari M, Annane D, Bauer M, Bellomo R, Bernard GR, Chiche JD, Coopersmith CM, Hotchkiss RS, Levy MM, Marshall JC, et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA. 2016; 315:801–10. https://doi.org/10.1001/jama.2016.0287 [PubMed]
- 2. Tan B, Wong JJ, Sultana R, Koh JC, Jit M, Mok YH, Lee JH. Global Case-Fatality Rates in Pediatric Severe Sepsis and Septic Shock: A Systematic Review and Meta-analysis. JAMA Pediatr. 2019; 173:352–62. https://doi.org/10.1001/jamapediatrics.2018.4839 [PubMed]
- 3. Li S, Wu H, Han D, Ma S, Fan W, Wang Y, Zhang R, Fan M, Huang Y, Fu X, Cao F. A Novel Mechanism of Mesenchymal Stromal Cell-Mediated Protection against Sepsis: Restricting Inflammasome Activation in Macrophages by Increasing Mitophagy and Decreasing Mitochondrial ROS. Oxid Med Cell Longev. 2018; 2018:3537609. https://doi.org/10.1155/2018/3537609 [PubMed]
- 4. Vandewalle J, Libert C. Sepsis: a failing starvation response. Trends Endocrinol Metab. 2022; 33:292–304. https://doi.org/10.1016/j.tem.2022.01.006 [PubMed]
- 5. Rudd KE, Johnson SC, Agesa KM, Shackelford KA, Tsoi D, Kievlan DR, Colombara DV, Ikuta KS, Kissoon N, Finfer S, Fleischmann-Struzek C, Machado FR, Reinhart KK, et al. Global, regional, and national sepsis incidence and mortality, 1990-2017: analysis for the Global Burden of Disease Study. Lancet. 2020; 395:200–11. https://doi.org/10.1016/S0140-6736(19)32989-7 [PubMed]
- 6. Lelubre C, Vincent JL. Mechanisms and treatment of organ failure in sepsis. Nat Rev Nephrol. 2018; 14:417–27. https://doi.org/10.1038/s41581-018-0005-7 [PubMed]
- 7. Chen XS, Cui JR, Meng XL, Wang SH, Wei W, Gao YL, Shou ST, Liu YC, Chai YF. Angiotensin-(1-7) ameliorates sepsis-induced cardiomyopathy by alleviating inflammatory response and mitochondrial damage through the NF-κB and MAPK pathways. J Transl Med. 2023; 21:2. https://doi.org/10.1186/s12967-022-03842-5 [PubMed]
- 8. Zhang WX, He BM, Wu Y, Qiao JF, Peng ZY. Melatonin protects against sepsis-induced cardiac dysfunction by regulating apoptosis and autophagy via activation of SIRT1 in mice. Life Sci. 2019; 217:8–15. https://doi.org/10.1016/j.lfs.2018.11.055 [PubMed]
- 9. Kuwabara S, Goggins E, Okusa MD. The Pathophysiology of Sepsis-Associated AKI. Clin J Am Soc Nephrol. 2022; 17:1050–69. https://doi.org/10.2215/CJN.00850122 [PubMed]
- 10. Bi CF, Liu J, Yang LS, Zhang JF. Research Progress on the Mechanism of Sepsis Induced Myocardial Injury. J Inflamm Res. 2022; 15:4275–90. https://doi.org/10.2147/JIR.S374117 [PubMed]
- 11. Papafilippou L, Claxton A, Dark P, Kostarelos K, Hadjidemetriou M. Nanotools for Sepsis Diagnosis and Treatment. Adv Healthc Mater. 2021; 10:e2001378. https://doi.org/10.1002/adhm.202001378 [PubMed]
- 12. Font MD, Thyagarajan B, Khanna AK. Sepsis and Septic Shock - Basics of diagnosis, pathophysiology and clinical decision making. Med Clin North Am. 2020; 104:573–85. https://doi.org/10.1016/j.mcna.2020.02.011 [PubMed]
- 13. Wang Y, Xu M, Yue P, Zhang D, Tong J, Li Y. Novel Insights Into the Potential Mechanisms of N6-Methyladenosine RNA Modification on Sepsis-Induced Cardiovascular Dysfunction: An Update Summary on Direct and Indirect Evidences. Front Cell Dev Biol. 2021; 9:772921. https://doi.org/10.3389/fcell.2021.772921 [PubMed]
- 14. Jia G, Fu Y, Zhao X, Dai Q, Zheng G, Yang Y, Yi C, Lindahl T, Pan T, Yang YG, He C. N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat Chem Biol. 2011; 7:885–7. https://doi.org/10.1038/nchembio.687 [PubMed]
- 15. Dominissini D, Moshitch-Moshkovitz S, Schwartz S, Salmon-Divon M, Ungar L, Osenberg S, Cesarkas K, Jacob-Hirsch J, Amariglio N, Kupiec M, Sorek R, Rechavi G. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature. 2012; 485:201–6. https://doi.org/10.1038/nature11112 [PubMed]
- 16. Meng Y, Zhang Q, Wang K, Zhang X, Yang R, Bi K, Chen W, Diao H. RBM15-mediated N6-methyladenosine modification affects COVID-19 severity by regulating the expression of multitarget genes. Cell Death Dis. 2021; 12:732. https://doi.org/10.1038/s41419-021-04012-z [PubMed]
- 17. Liu J, Sun G, Pan S, Qin M, Ouyang R, Li Z, Huang J. The Cancer Genome Atlas (TCGA) based m6A methylation-related genes predict prognosis in hepatocellular carcinoma. Bioengineered. 2020; 11:759–68. https://doi.org/10.1080/21655979.2020.1787764 [PubMed]
- 18. Xu H, Lin C, Yang J, Chen X, Chen Y, Chen J, Guo A, Hu C. The Role of N6-Methyladenosine in Inflammatory Diseases. Oxid Med Cell Longev. 2022; 2022:9744771. https://doi.org/10.1155/2022/9744771 [PubMed]
- 19. Wang X, Ding Y, Li R, Zhang R, Ge X, Gao R, Wang M, Huang Y, Zhang F, Zhao B, Liao W, Du J. N6-methyladenosine of Spi2a attenuates inflammation and sepsis-associated myocardial dysfunction in mice. Nat Commun. 2023; 14:1185. https://doi.org/10.1038/s41467-023-36865-7 [PubMed]
- 20. Lin C, Chen ST, Chien SY, Kuo SJ, Chen DR. Use of high-dose nandrolone aggravates septic shock in a mouse model. Kaohsiung J Med Sci. 2011; 27:222–9. https://doi.org/10.1016/j.kjms.2010.12.015 [PubMed]
- 21. Shen ZJ, Han YC, Nie MW, Wang YN, Xiang RL, Xie HZ. Genome-wide identification of altered RNA m6A profiles in vascular tissue of septic rats. Aging (Albany NY). 2021; 13:21610–27. https://doi.org/10.18632/aging.203506 [PubMed]
- 22. Levine B, Kroemer G. Biological Functions of Autophagy Genes: A Disease Perspective. Cell. 2019; 176:11–42. https://doi.org/10.1016/j.cell.2018.09.048 [PubMed]
- 23. Ho J, Yu J, Wong SH, Zhang L, Liu X, Wong WT, Leung CC, Choi G, Wang MH, Gin T, Chan MT, Wu WK. Autophagy in sepsis: Degradation into exhaustion? Autophagy. 2016; 12:1073–82. https://doi.org/10.1080/15548627.2016.1179410 [PubMed]
- 24. Deng Z, Sun M, Wu J, Fang H, Cai S, An S, Huang Q, Chen Z, Wu C, Zhou Z, Hu H, Zeng Z. SIRT1 attenuates sepsis-induced acute kidney injury via Beclin1 deacetylation-mediated autophagy activation. Cell Death Dis. 2021; 12:217. https://doi.org/10.1038/s41419-021-03508-y [PubMed]
- 25. Sun Y, Yao X, Zhang QJ, Zhu M, Liu ZP, Ci B, Xie Y, Carlson D, Rothermel BA, Sun Y, Levine B, Hill JA, Wolf SE, et al. Beclin-1-Dependent Autophagy Protects the Heart During Sepsis. Circulation. 2018; 138:2247–62. https://doi.org/10.1161/CIRCULATIONAHA.117.032821 [PubMed]
- 26. Chen H, Lin H, Dong B, Wang Y, Yu Y, Xie K. Hydrogen alleviates cell damage and acute lung injury in sepsis via PINK1/Parkin-mediated mitophagy. Inflamm Res. 2021; 70:915–30. https://doi.org/10.1007/s00011-021-01481-y [PubMed]
- 27. Wang F, Liao Y, Zhang M, Zhu Y, Wang W, Cai H, Liang J, Song F, Hou C, Huang S, Zhang Y, Wang C, Hou J. N6-methyladenosine demethyltransferase FTO-mediated autophagy in malignant development of oral squamous cell carcinoma. Oncogene. 2021; 40:3885–98. https://doi.org/10.1038/s41388-021-01820-7 [PubMed]
- 28. Lv D, Gimple RC, Zhong C, Wu Q, Yang K, Prager BC, Godugu B, Qiu Z, Zhao L, Zhang G, Dixit D, Lee D, Shen JZ, et al. PDGF signaling inhibits mitophagy in glioblastoma stem cells through N6-methyladenosine. Dev Cell. 2022; 57:1466–81.e6. https://doi.org/10.1016/j.devcel.2022.05.007 [PubMed]
- 29. Liu L, Li H, Hu D, Wang Y, Shao W, Zhong J, Yang S, Liu J, Zhang J. Insights into N6-methyladenosine and programmed cell death in cancer. Mol Cancer. 2022; 21:32. https://doi.org/10.1186/s12943-022-01508-w [PubMed]
- 30. Zhang S, Liu F, Wu Z, Xie J, Yang Y, Qiu H. Contribution of m6A subtype classification on heterogeneity of sepsis. Ann Transl Med. 2020; 8:306. https://doi.org/10.21037/atm.2020.03.07 [PubMed]
- 31. Fu Y, Dominissini D, Rechavi G, He C. Gene expression regulation mediated through reversible m6A RNA methylation. Nat Rev Genet. 2014; 15:293–306. https://doi.org/10.1038/nrg3724 [PubMed]
- 32. Meyer KD, Saletore Y, Zumbo P, Elemento O, Mason CE, Jaffrey SR. Comprehensive analysis of mRNA methylation reveals enrichment in 3' UTRs and near stop codons. Cell. 2012; 149:1635–46. https://doi.org/10.1016/j.cell.2012.05.003 [PubMed]
- 33. Perry RP, Kelley DE. Methylated constituents of heterogeneous nuclear RNA: presence in blocked 5' terminal structures. Cell. 1975; 6:13–9. https://doi.org/10.1016/0092-8674(75)90068-9 [PubMed]
- 34. Perry RP, Kelley DE, Friderici K, Rottman F. The methylated constituents of L cell messenger RNA: evidence for an unusual cluster at the 5' terminus. Cell. 1975; 4:387–94. https://doi.org/10.1016/0092-8674(75)90159-2 [PubMed]
- 35. Meyer KD, Patil DP, Zhou J, Zinoviev A, Skabkin MA, Elemento O, Pestova TV, Qian SB, Jaffrey SR. 5' UTR m(6)A Promotes Cap-Independent Translation. Cell. 2015; 163:999–1010. https://doi.org/10.1016/j.cell.2015.10.012 [PubMed]
- 36. Berulava T, Buchholz E, Elerdashvili V, Pena T, Islam MR, Lbik D, Mohamed BA, Renner A, von Lewinski D, Sacherer M, Bohnsack KE, Bohnsack MT, Jain G, et al. Changes in m6A RNA methylation contribute to heart failure progression by modulating translation. Eur J Heart Fail. 2020; 22:54–66. https://doi.org/10.1002/ejhf.1672 [PubMed]
- 37. Shi H, Wei J, He C. Where, When, and How: Context-Dependent Functions of RNA Methylation Writers, Readers, and Erasers. Mol Cell. 2019; 74:640–50. https://doi.org/10.1016/j.molcel.2019.04.025 [PubMed]
- 38. Doxtader KA, Wang P, Scarborough AM, Seo D, Conrad NK, Nam Y. Structural Basis for Regulation of METTL16, an S-Adenosylmethionine Homeostasis Factor. Mol Cell. 2018; 71:1001–11.e4. https://doi.org/10.1016/j.molcel.2018.07.025 [PubMed]
- 39. Han J, Kong H, Wang X, Zhang XA. Novel insights into the interaction between N6-methyladenosine methylation and noncoding RNAs in musculoskeletal disorders. Cell Prolif. 2022; 55:e13294. https://doi.org/10.1111/cpr.13294 [PubMed]
- 40. Barbieri I, Kouzarides T. Role of RNA modifications in cancer. Nat Rev Cancer. 2020; 20:303–22. https://doi.org/10.1038/s41568-020-0253-2 [PubMed]
- 41. An Y, Duan H. The role of m6A RNA methylation in cancer metabolism. Mol Cancer. 2022; 21:14. https://doi.org/10.1186/s12943-022-01500-4 [PubMed]
- 42. Liu N, Zhou KI, Parisien M, Dai Q, Diatchenko L, Pan T. N6-methyladenosine alters RNA structure to regulate binding of a low-complexity protein. Nucleic Acids Res. 2017; 45:6051–63. https://doi.org/10.1093/nar/gkx141 [PubMed]
- 43. Suo L, Liu C, Zhang QY, Yao MD, Ma Y, Yao J, Jiang Q, Yan B. METTL3-mediated N6-methyladenosine modification governs pericyte dysfunction during diabetes-induced retinal vascular complication. Theranostics. 2022; 12:277–89. https://doi.org/10.7150/thno.63441 [PubMed]
- 44. Liu L, He J, Sun G, Huang N, Bian Z, Xu C, Zhang Y, Cui Z, Xu W, Sun F, Zhuang C, Man Q, Gu S. The N6-methyladenosine modification enhances ferroptosis resistance through inhibiting SLC7A11 mRNA deadenylation in hepatoblastoma. Clin Transl Med. 2022; 12:e778. https://doi.org/10.1002/ctm2.778 [PubMed]
- 45. Xu S, Li Y, Chen JP, Li DZ, Jiang Q, Wu T, Zhou XZ. Oxygen glucose deprivation/re-oxygenation-induced neuronal cell death is associated with Lnc-D63785 m6A methylation and miR-422a accumulation. Cell Death Dis. 2020; 11:816. https://doi.org/10.1038/s41419-020-03021-8 [PubMed]
- 46. Wu Y, Chang N, Zhang Y, Zhang X, Xu L, Che Y, Qiao T, Wu B, Zhou Y, Jiang J, Xiong J, Zhang J, Zhang J. METTL3-mediated m6A mRNA modification of FBXW7 suppresses lung adenocarcinoma. J Exp Clin Cancer Res. 2021; 40:90. https://doi.org/10.1186/s13046-021-01880-3 [PubMed]
- 47. Yue B, Song C, Yang L, Cui R, Cheng X, Zhang Z, Zhao G. METTL3-mediated N6-methyladenosine modification is critical for epithelial-mesenchymal transition and metastasis of gastric cancer. Mol Cancer. 2019; 18:142. https://doi.org/10.1186/s12943-019-1065-4 [PubMed]
- 48. Gong R, Wang X, Li H, Liu S, Jiang Z, Zhao Y, Yu Y, Han Z, Yu Y, Dong C, Li S, Xu B, Zhang W, et al. Loss of m6A methyltransferase METTL3 promotes heart regeneration and repair after myocardial injury. Pharmacol Res. 2021; 174:105845. https://doi.org/10.1016/j.phrs.2021.105845 [PubMed]
- 49. Su Y, Xu R, Zhang R, Qu Y, Zuo W, Ji Z, Geng H, Pan M, Ma G. N6-methyladenosine methyltransferase plays a role in hypoxic preconditioning partially through the interaction with lncRNA H19. Acta Biochim Biophys Sin (Shanghai). 2020; 52:1306–15. https://doi.org/10.1093/abbs/gmaa130 [PubMed]
- 50. Cao Y, Wang Z, Yan Y, Ji L, He J, Xuan B, Shen C, Ma Y, Jiang S, Ma D, Tong T, Zhang X, Gao Z, et al. Enterotoxigenic Bacteroidesfragilis Promotes Intestinal Inflammation and Malignancy by Inhibiting Exosome-Packaged miR-149-3p. Gastroenterology. 2021; 161:1552–66.e12. https://doi.org/10.1053/j.gastro.2021.08.003 [PubMed]
- 51. Chen X, Xu M, Xu X, Zeng K, Liu X, Pan B, Li C, Sun L, Qin J, Xu T, He B, Pan Y, Sun H, Wang S. METTL14-mediated N6-methyladenosine modification of SOX4 mRNA inhibits tumor metastasis in colorectal cancer. Mol Cancer. 2020; 19:106. https://doi.org/10.1186/s12943-020-01220-7 [PubMed]
- 52. Yang X, Zhang S, He C, Xue P, Zhang L, He Z, Zang L, Feng B, Sun J, Zheng M. METTL14 suppresses proliferation and metastasis of colorectal cancer by down-regulating oncogenic long non-coding RNA XIST. Mol Cancer. 2020; 19:46. https://doi.org/10.1186/s12943-020-1146-4 [PubMed]
- 53. Xie Q, Li Z, Luo X, Wang D, Zhou Y, Zhao J, Gao S, Yang Y, Fu W, Kong L, Sun T. piRNA-14633 promotes cervical cancer cell malignancy in a METTL14-dependent m6A RNA methylation manner. J Transl Med. 2022; 20:51. https://doi.org/10.1186/s12967-022-03257-2 [PubMed]
- 54. Meng L, Lin H, Huang X, Weng J, Peng F, Wu S. METTL14 suppresses pyroptosis and diabetic cardiomyopathy by downregulating TINCR lncRNA. Cell Death Dis. 2022; 13:38. https://doi.org/10.1038/s41419-021-04484-z [PubMed]
- 55. Dai YZ, Liu YD, Li J, Chen MT, Huang M, Wang F, Yang QS, Yuan JH, Sun SH. METTL16 promotes hepatocellular carcinoma progression through downregulating RAB11B-AS1 in an m6A-dependent manner. Cell Mol Biol Lett. 2022; 27:41. https://doi.org/10.1186/s11658-022-00342-8 [PubMed]
- 56. Wang XK, Zhang YW, Wang CM, Li B, Zhang TZ, Zhou WJ, Cheng LJ, Huo MY, Zhang CH, He YL. METTL16 promotes cell proliferation by up-regulating cyclin D1 expression in gastric cancer. J Cell Mol Med. 2021; 25:6602–17. https://doi.org/10.1111/jcmm.16664 [PubMed]
- 57. Cho S, Lee G, Pickering BF, Jang C, Park JH, He L, Mathur L, Kim SS, Jung S, Tang HW, Monette S, Rabinowitz JD, Perrimon N, et al. mTORC1 promotes cell growth via m6A-dependent mRNA degradation. Mol Cell. 2021; 81:2064–75.e8. https://doi.org/10.1016/j.molcel.2021.03.010 [PubMed]
- 58. Chen Y, Peng C, Chen J, Chen D, Yang B, He B, Hu W, Zhang Y, Liu H, Dai L, Xie H, Zhou L, Wu J, Zheng S. WTAP facilitates progression of hepatocellular carcinoma via m6A-HuR-dependent epigenetic silencing of ETS1. Mol Cancer. 2019; 18:127. https://doi.org/10.1186/s12943-019-1053-8 [PubMed]
- 59. Lan T, Li H, Zhang D, Xu L, Liu H, Hao X, Yan X, Liao H, Chen X, Xie K, Li J, Liao M, Huang J, et al. KIAA1429 contributes to liver cancer progression through N6-methyladenosine-dependent post-transcriptional modification of GATA3. Mol Cancer. 2019; 18:186. https://doi.org/10.1186/s12943-019-1106-z [PubMed]
- 60. Zhou Y, Pei Z, Maimaiti A, Zheng L, Zhu Z, Tian M, Zhou Z, Tan F, Pei Q, Li Y, Liu W. m6A methyltransferase KIAA1429 acts as an oncogenic factor in colorectal cancer by regulating SIRT1 in an m6A-dependent manner. Cell Death Discov. 2022; 8:83. https://doi.org/10.1038/s41420-022-00878-w [PubMed]
- 61. Zhang C, Sun Q, Zhang X, Qin N, Pu Z, Gu Y, Yan C, Zhu M, Dai J, Wang C, Li N, Jin G, Ma H, et al. Gene amplification-driven RNA methyltransferase KIAA1429 promotes tumorigenesis by regulating BTG2 via m6A-YTHDF2-dependent in lung adenocarcinoma. Cancer Commun (Lond). 2022; 42:609–26. https://doi.org/10.1002/cac2.12325 [PubMed]
- 62. Barros-Silva D, Lobo J, Guimarães-Teixeira C, Carneiro I, Oliveira J, Martens-Uzunova ES, Henrique R, Jerónimo C. VIRMA-Dependent N6-Methyladenosine Modifications Regulate the Expression of Long Non-Coding RNAs CCAT1 and CCAT2 in Prostate Cancer. Cancers (Basel). 2020; 12:771. https://doi.org/10.3390/cancers12040771 [PubMed]
- 63. Wang X, Tian L, Li Y, Wang J, Yan B, Yang L, Li Q, Zhao R, Liu M, Wang P, Sun Y. RBM15 facilitates laryngeal squamous cell carcinoma progression by regulating TMBIM6 stability through IGF2BP3 dependent. J Exp Clin Cancer Res. 2021; 40:80. https://doi.org/10.1186/s13046-021-01871-4 [PubMed]
- 64. Yang F, Liu Y, Xiao J, Li B, Chen Y, Hu A, Zeng J, Liu Z, Liu H. Circ-CTNNB1 drives aerobic glycolysis and osteosarcoma progression via m6A modification through interacting with RBM15. Cell Prolif. 2023; 56:e13344. https://doi.org/10.1111/cpr.13344 [PubMed]
- 65. Fang J, Wu X, He J, Zhang H, Chen X, Zhang H, Novakovic B, Qi H, Yu X. RBM15 suppresses hepatic insulin sensitivity of offspring of gestational diabetes mellitus mice via m6A-mediated regulation of CLDN4. Mol Med. 2023; 29:23. https://doi.org/10.1186/s10020-023-00615-8 [PubMed]
- 66. Wang J, Yang Y, Sun F, Luo Y, Yang Y, Li J, Hu W, Tao H, Lu C, Yang JJ. ALKBH5 attenuates mitochondrial fission and ameliorates liver fibrosis by reducing Drp1 methylation. Pharmacol Res. 2023; 187:106608. https://doi.org/10.1016/j.phrs.2022.106608 [PubMed]
- 67. Yang Z, Cai Z, Yang C, Luo Z, Bao X. ALKBH5 regulates STAT3 activity to affect the proliferation and tumorigenicity of osteosarcoma via an m6A-YTHDF2-dependent manner. EBioMedicine. 2022; 80:104019. https://doi.org/10.1016/j.ebiom.2022.104019 [PubMed]
- 68. Tang B, Yang Y, Kang M, Wang Y, Wang Y, Bi Y, He S, Shimamoto F. m6A demethylase ALKBH5 inhibits pancreatic cancer tumorigenesis by decreasing WIF-1 RNA methylation and mediating Wnt signaling. Mol Cancer. 2020; 19:3. https://doi.org/10.1186/s12943-019-1128-6 [PubMed]
- 69. Chen J, Xu C, Yang K, Gao R, Cao Y, Liang L, Chen S, Xu S, Rong R, Wang J, Zhu T. Inhibition of ALKBH5 attenuates I/R-induced renal injury in male mice by promoting Ccl28 m6A modification and increasing Treg recruitment. Nat Commun. 2023; 14:1161. https://doi.org/10.1038/s41467-023-36747-y [PubMed]
- 70. Mathiyalagan P, Adamiak M, Mayourian J, Sassi Y, Liang Y, Agarwal N, Jha D, Zhang S, Kohlbrenner E, Chepurko E, Chen J, Trivieri MG, Singh R, et al. FTO-Dependent N6-Methyladenosine Regulates Cardiac Function During Remodeling and Repair. Circulation. 2019; 139:51–32. https://doi.org/10.1161/CIRCULATIONAHA.118.033794 [PubMed]
- 71. Jiang ZX, Wang YN, Li ZY, Dai ZH, He Y, Chu K, Gu JY, Ji YX, Sun NX, Yang F, Li W. The m6A mRNA demethylase FTO in granulosa cells retards FOS-dependent ovarian aging. Cell Death Dis. 2021; 12:744. https://doi.org/10.1038/s41419-021-04016-9 [PubMed]
- 72. Wei K, Gao Y, Wang B, Qu YX. Methylation recognition protein YTH N6-methyladenosine RNA binding protein 1 (YTHDF1) regulates the proliferation, migration and invasion of osteosarcoma by regulating m6A level of CCR4-NOT transcription complex subunit 7 (CNOT7). Bioengineered. 2022; 13:5236–50. https://doi.org/10.1080/21655979.2022.2037381 [PubMed]
- 73. Li J, Xie H, Ying Y, Chen H, Yan H, He L, Xu M, Xu X, Liang Z, Liu B, Wang X, Zheng X, Xie L. YTHDF2 mediates the mRNA degradation of the tumor suppressors to induce AKT phosphorylation in N6-methyladenosine-dependent way in prostate cancer. Mol Cancer. 2020; 19:152. https://doi.org/10.1186/s12943-020-01267-6 [PubMed]
- 74. Chai RC, Chang YZ, Chang X, Pang B, An SY, Zhang KN, Chang YH, Jiang T, Wang YZ. YTHDF2 facilitates UBXN1 mRNA decay by recognizing METTL3-mediated m6A modification to activate NF-κB and promote the malignant progression of glioma. J Hematol Oncol. 2021; 14:109. https://doi.org/10.1186/s13045-021-01124-z [PubMed]
- 75. Xu H, Wang Z, Chen M, Zhao W, Tao T, Ma L, Ni Y, Li W. YTHDF2 alleviates cardiac hypertrophy via regulating Myh7 mRNA decoy. Cell Biosci. 2021; 11:132. https://doi.org/10.1186/s13578-021-00649-7 [PubMed]
- 76. Liang D, Lin WJ, Ren M, Qiu J, Yang C, Wang X, Li N, Zeng T, Sun K, You L, Yan L, Wang W. m6A reader YTHDC1 modulates autophagy by targeting SQSTM1 in diabetic skin. Autophagy. 2022; 18:1318–37. https://doi.org/10.1080/15548627.2021.1974175 [PubMed]
- 77. Yu D, Pan M, Li Y, Lu T, Wang Z, Liu C, Hu G. RNA N6-methyladenosine reader IGF2BP2 promotes lymphatic metastasis and epithelial-mesenchymal transition of head and neck squamous carcinoma cells via stabilizing slug mRNA in an m6A-dependent manner. J Exp Clin Cancer Res. 2022; 41:6. https://doi.org/10.1186/s13046-021-02212-1 [PubMed]
- 78. Wang JN, Wang F, Ke J, Li Z, Xu CH, Yang Q, Chen X, He XY, He Y, Suo XG, Li C, Yu JT, Jiang L, et al. Inhibition of METTL3 attenuates renal injury and inflammation by alleviating TAB3 m6A modifications via IGF2BP2-dependent mechanisms. Sci Transl Med. 2022; 14:eabk2709. https://doi.org/10.1126/scitranslmed.abk2709 [PubMed]
- 79. Jiang L, Liu X, Hu X, Gao L, Zeng H, Wang X, Huang Y, Zhu W, Wang J, Wen J, Meng X, Wu Y. METTL3-mediated m6A modification of TIMP2 mRNA promotes podocyte injury in diabetic nephropathy. Mol Ther. 2022; 30:1721–40. https://doi.org/10.1016/j.ymthe.2022.01.002 [PubMed]
- 80. Ban Y, Tan P, Cai J, Li J, Hu M, Zhou Y, Mei Y, Tan Y, Li X, Zeng Z, Xiong W, Li G, Li X, et al. LNCAROD is stabilized by m6A methylation and promotes cancer progression via forming a ternary complex with HSPA1A and YBX1 in head and neck squamous cell carcinoma. Mol Oncol. 2020; 14:1282–96. https://doi.org/10.1002/1878-0261.12676 [PubMed]
- 81. Wang X, Feng J, Xue Y, Guan Z, Zhang D, Liu Z, Gong Z, Wang Q, Huang J, Tang C, Zou T, Yin P. Structural basis of N(6)-adenosine methylation by the METTL3-METTL14 complex. Nature. 2016; 534:575–8. https://doi.org/10.1038/nature18298 [PubMed]
- 82. Du Y, Hou G, Zhang H, Dou J, He J, Guo Y, Li L, Chen R, Wang Y, Deng R, Huang J, Jiang B, Xu M, et al. SUMOylation of the m6A-RNA methyltransferase METTL3 modulates its function. Nucleic Acids Res. 2018; 46:5195–208. https://doi.org/10.1093/nar/gky156 [PubMed]
- 83. Liu J, Yue Y, Han D, Wang X, Fu Y, Zhang L, Jia G, Yu M, Lu Z, Deng X, Dai Q, Chen W, He C. A METTL3-METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat Chem Biol. 2014; 10:93–5. https://doi.org/10.1038/nchembio.1432 [PubMed]
- 84. Huang J, Dong X, Gong Z, Qin LY, Yang S, Zhu YL, Wang X, Zhang D, Zou T, Yin P, Tang C. Solution structure of the RNA recognition domain of METTL3-METTL14 N6-methyladenosine methyltransferase. Protein Cell. 2019; 10:272–84. https://doi.org/10.1007/s13238-018-0518-7 [PubMed]
- 85. Ping XL, Sun BF, Wang L, Xiao W, Yang X, Wang WJ, Adhikari S, Shi Y, Lv Y, Chen YS, Zhao X, Li A, Yang Y, et al. Mammalian WTAP is a regulatory subunit of the RNA N6-methyladenosine methyltransferase. Cell Res. 2014; 24:177–89. https://doi.org/10.1038/cr.2014.3 [PubMed]
- 86. Villa E, Sahu U, O’Hara BP, Ali ES, Helmin KA, Asara JM, Gao P, Singer BD, Ben-Sahra I. mTORC1 stimulates cell growth through SAM synthesis and m6A mRNA-dependent control of protein synthesis. Mol Cell. 2021; 81:2076–93.e9. https://doi.org/10.1016/j.molcel.2021.03.009 [PubMed]
- 87. Mendel M, Chen KM, Homolka D, Gos P, Pandey RR, McCarthy AA, Pillai RS. Methylation of Structured RNA by the m6A Writer METTL16 Is Essential for Mouse Embryonic Development. Mol Cell. 2018; 71:986–1000.e11. https://doi.org/10.1016/j.molcel.2018.08.004 [PubMed]
- 88. Pendleton KE, Chen B, Liu K, Hunter OV, Xie Y, Tu BP, Conrad NK. The U6 snRNA m6A Methyltransferase METTL16 Regulates SAM Synthetase Intron Retention. Cell. 2017; 169:824–35.e14. https://doi.org/10.1016/j.cell.2017.05.003 [PubMed]
- 89. Wang F, Zhang J, Lin X, Yang L, Zhou Q, Mi X, Li Q, Wang S, Li D, Liu XM, Zhou J. METTL16 promotes translation and lung tumorigenesis by sequestering cytoplasmic eIF4E2. Cell Rep. 2023; 42:112150. https://doi.org/10.1016/j.celrep.2023.112150 [PubMed]
- 90. Su S, Li S, Deng T, Gao M, Yin Y, Wu B, Peng C, Liu J, Ma J, Zhang K. Cryo-EM structures of human m6A writer complexes. Cell Res. 2022; 32:982–94. https://doi.org/10.1038/s41422-022-00725-8 [PubMed]
- 91. Yue Y, Liu J, Cui X, Cao J, Luo G, Zhang Z, Cheng T, Gao M, Shu X, Ma H, Wang F, Wang X, Shen B, et al. VIRMA mediates preferential m6A mRNA methylation in 3'UTR and near stop codon and associates with alternative polyadenylation. Cell Discov. 2018; 4:10. https://doi.org/10.1038/s41421-018-0019-0 [PubMed]
- 92. Patil DP, Chen CK, Pickering BF, Chow A, Jackson C, Guttman M, Jaffrey SR. m(6)A RNA methylation promotes XIST-mediated transcriptional repression. Nature. 2016; 537:369–73. https://doi.org/10.1038/nature19342 [PubMed]
- 93. Ma H, Wang X, Cai J, Dai Q, Natchiar SK, Lv R, Chen K, Lu Z, Chen H, Shi YG, Lan F, Fan J, Klaholz BP, Pan T, Shi Y, He C. N6-Methyladenosine methyltransferase ZCCHC4 mediates ribosomal RNA methylation. Nat Chem Biol. 2019; 15:88–94. https://doi.org/10.1038/s41589-018-0184-3 [PubMed]
- 94. Pinto R, Vågbø CB, Jakobsson ME, Kim Y, Baltissen MP, O’Donohue MF, Guzmán UH, Małecki JM, Wu J, Kirpekar F, Olsen JV, Gleizes PE, Vermeulen M, et al. The human methyltransferase ZCCHC4 catalyses N6-methyladenosine modification of 28S ribosomal RNA. Nucleic Acids Res. 2020; 48:830–46. https://doi.org/10.1093/nar/gkz1147 [PubMed]
- 95. Ren W, Lu J, Huang M, Gao L, Li D, Wang GG, Song J. Structure and regulation of ZCCHC4 in m6A-methylation of 28S rRNA. Nat Commun. 2019; 10:5042. https://doi.org/10.1038/s41467-019-12923-x [PubMed]
- 96. Bartosovic M, Molares HC, Gregorova P, Hrossova D, Kudla G, Vanacova S. N6-methyladenosine demethylase FTO targets pre-mRNAs and regulates alternative splicing and 3'-end processing. Nucleic Acids Res. 2017; 45:11356–70. https://doi.org/10.1093/nar/gkx778 [PubMed]
- 97. Yu J, Chen M, Huang H, Zhu J, Song H, Zhu J, Park J, Ji SJ. Dynamic m6A modification regulates local translation of mRNA in axons. Nucleic Acids Res. 2018; 46:1412–23. https://doi.org/10.1093/nar/gkx1182 [PubMed]
- 98. Aik W, Scotti JS, Choi H, Gong L, Demetriades M, Schofield CJ, McDonough MA. Structure of human RNA N6-methyladenine demethylase ALKBH5 provides insights into its mechanisms of nucleic acid recognition and demethylation. Nucleic Acids Res. 2014; 42:4741–54. https://doi.org/10.1093/nar/gku085 [PubMed]
- 99. Zheng G, Dahl JA, Niu Y, Fedorcsak P, Huang CM, Li CJ, Vågbø CB, Shi Y, Wang WL, Song SH, Lu Z, Bosmans RP, Dai Q, et al. ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility. Mol Cell. 2013; 49:18–29. https://doi.org/10.1016/j.molcel.2012.10.015 [PubMed]
- 100. Liu Y, Zhou T, Wang Q, Fu R, Zhang Z, Chen N, Li Z, Gao G, Peng S, Yang D. m6 A demethylase ALKBH5 drives denervation-induced muscle atrophy by targeting HDAC4 to activate FoxO3 signalling. J Cachexia Sarcopenia Muscle. 2022; 13:1210–23. https://doi.org/10.1002/jcsm.12929 [PubMed]
- 101. Yu T, Yao L, Yin H, Teng Y, Hong M, Wu Q. ALKBH5 Promotes Multiple Myeloma Tumorigenicity through inducing m6A-demethylation of SAV1 mRNA and Myeloma Stem Cell Phenotype. Int J Biol Sci. 2022; 18:2235–48. https://doi.org/10.7150/ijbs.64943 [PubMed]
- 102. Han Z, Wang X, Xu Z, Cao Y, Gong R, Yu Y, Yu Y, Guo X, Liu S, Yu M, Ma W, Zhao Y, Xu J, et al. ALKBH5 regulates cardiomyocyte proliferation and heart regeneration by demethylating the mRNA of YTHDF1. Theranostics. 2021; 11:3000–16. https://doi.org/10.7150/thno.47354 [PubMed]
- 103. Koh CWQ, Goh YT, Goh WSS. Atlas of quantitative single-base-resolution N6-methyl-adenine methylomes. Nat Commun. 2019; 10:5636. https://doi.org/10.1038/s41467-019-13561-z [PubMed]
- 104. Zhou KI, Pan T. An additional class of m6A readers. Nat Cell Biol. 2018; 20:230–2. https://doi.org/10.1038/s41556-018-0046-y [PubMed]
- 105. Wang X, Zhao BS, Roundtree IA, Lu Z, Han D, Ma H, Weng X, Chen K, Shi H, He C. N(6)-methyladenosine Modulates Messenger RNA Translation Efficiency. Cell. 2015; 161:1388–99. https://doi.org/10.1016/j.cell.2015.05.014 [PubMed]
- 106. Wang X, Lu Z, Gomez A, Hon GC, Yue Y, Han D, Fu Y, Parisien M, Dai Q, Jia G, Ren B, Pan T, He C. N6-methyladenosine-dependent regulation of messenger RNA stability. Nature. 2014; 505:117–20. https://doi.org/10.1038/nature12730 [PubMed]
- 107. Shi H, Wang X, Lu Z, Zhao BS, Ma H, Hsu PJ, Liu C, He C. YTHDF3 facilitates translation and decay of N6-methyladenosine-modified RNA. Cell Res. 2017; 27:315–28. https://doi.org/10.1038/cr.2017.15 [PubMed]
- 108. Zaccara S, Jaffrey SR. A Unified Model for the Function of YTHDF Proteins in Regulating m6A-Modified mRNA. Cell. 2020; 181:1582–95.e18. https://doi.org/10.1016/j.cell.2020.05.012 [PubMed]
- 109. Louloupi A, Ntini E, Conrad T, Ørom UA. Transient N-6-Methyladenosine Transcriptome Sequencing Reveals a Regulatory Role of m6A in Splicing Efficiency. Cell Rep. 2018; 23:3429–37. https://doi.org/10.1016/j.celrep.2018.05.077 [PubMed]
- 110. Xiao W, Adhikari S, Dahal U, Chen YS, Hao YJ, Sun BF, Sun HY, Li A, Ping XL, Lai WY, Wang X, Ma HL, Huang CM, et al. Nuclear m(6)A Reader YTHDC1 Regulates mRNA Splicing. Mol Cell. 2016; 61:507–19. https://doi.org/10.1016/j.molcel.2016.01.012 [PubMed]
- 111. Hsu PJ, Zhu Y, Ma H, Guo Y, Shi X, Liu Y, Qi M, Lu Z, Shi H, Wang J, Cheng Y, Luo G, Dai Q, et al. Ythdc2 is an N6-methyladenosine binding protein that regulates mammalian spermatogenesis. Cell Res. 2017; 27:1115–27. https://doi.org/10.1038/cr.2017.99 [PubMed]
- 112. Huang H, Weng H, Sun W, Qin X, Shi H, Wu H, Zhao BS, Mesquita A, Liu C, Yuan CL, Hu YC, Hüttelmaier S, Skibbe JR, et al. Recognition of RNA N6-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat Cell Biol. 2018; 20:285–95. https://doi.org/10.1038/s41556-018-0045-z [PubMed]
- 113. Johnson AG, Petrov AN, Fuchs G, Majzoub K, Grosely R, Choi J, Puglisi JD. Fluorescently-tagged human eIF3 for single-molecule spectroscopy. Nucleic Acids Res. 2018; 46:e8. https://doi.org/10.1093/nar/gkx1050 [PubMed]
- 114. Li T, Hu PS, Zuo Z, Lin JF, Li X, Wu QN, Chen ZH, Zeng ZL, Wang F, Zheng J, Chen D, Li B, Kang TB, Xie D, Lin D, Ju HQ, Xu RH. METTL3 facilitates tumor progression via an m6A-IGF2BP2-dependent mechanism in colorectal carcinoma. Mol Cancer. 2019; 18:112. https://doi.org/10.1186/s12943-019-1038-7 [PubMed]
- 115. Dorn LE, Lasman L, Chen J, Xu X, Hund TJ, Medvedovic M, Hanna JH, van Berlo JH, Accornero F. The N6-Methyladenosine mRNA Methylase METTL3 Controls Cardiac Homeostasis and Hypertrophy. Circulation. 2019; 139:533–45. https://doi.org/10.1161/CIRCULATIONAHA.118.036146 [PubMed]
- 116. Zhao T, Wang J, Wu Y, Han L, Chen J, Wei Y, Shen L, Long C, Wu S, Wei G. Increased m6A modification of RNA methylation related to the inhibition of demethylase FTO contributes to MEHP-induced Leydig cell injury☆. Environ Pollut. 2021; 268:115627. https://doi.org/10.1016/j.envpol.2020.115627 [PubMed]
- 117. Yin H, Zhang X, Yang P, Zhang X, Peng Y, Li D, Yu Y, Wu Y, Wang Y, Zhang J, Ding X, Wang X, Yang A, Zhang R. RNA m6A methylation orchestrates cancer growth and metastasis via macrophage reprogramming. Nat Commun. 2021; 12:1394. https://doi.org/10.1038/s41467-021-21514-8 [PubMed]
- 118. Choe J, Lin S, Zhang W, Liu Q, Wang L, Ramirez-Moya J, Du P, Kim W, Tang S, Sliz P, Santisteban P, George RE, Richards WG, et al. mRNA circularization by METTL3-eIF3h enhances translation and promotes oncogenesis. Nature. 2018; 561:556–60. https://doi.org/10.1038/s41586-018-0538-8 [PubMed]
- 119. Gao XQ, Zhang YH, Liu F, Ponnusamy M, Zhao XM, Zhou LY, Zhai M, Liu CY, Li XM, Wang M, Shan C, Shan PP, Wang Y, et al. The piRNA CHAPIR regulates cardiac hypertrophy by controlling METTL3-dependent N6-methyladenosine methylation of Parp10 mRNA. Nat Cell Biol. 2020; 22:1319–31. https://doi.org/10.1038/s41556-020-0576-y [PubMed]
- 120. Xu Y, Song M, Hong Z, Chen W, Zhang Q, Zhou J, Yang C, He Z, Yu J, Peng X, Zhu Q, Li S, Ji K, et al. The N6-methyladenosine METTL3 regulates tumorigenesis and glycolysis by mediating m6A methylation of the tumor suppressor LATS1 in breast cancer. J Exp Clin Cancer Res. 2023; 42:10. https://doi.org/10.1186/s13046-022-02581-1 [PubMed]
- 121. Xu Z, Qin Y, Lv B, Tian Z, Zhang B. Intermittent Fasting Improves High-Fat Diet-Induced Obesity Cardiomyopathy via Alleviating Lipid Deposition and Apoptosis and Decreasing m6A Methylation in the Heart. Nutrients. 2022; 14:251. https://doi.org/10.3390/nu14020251 [PubMed]
- 122. Chen Y, Zhao Y, Chen J, Peng C, Zhang Y, Tong R, Cheng Q, Yang B, Feng X, Lu Y, Xie H, Zhou L, Wu J, Zheng S. ALKBH5 suppresses malignancy of hepatocellular carcinoma via m6A-guided epigenetic inhibition of LYPD1. Mol Cancer. 2020; 19:123. https://doi.org/10.1186/s12943-020-01239-w [PubMed]
- 123. Guo X, Li K, Jiang W, Hu Y, Xiao W, Huang Y, Feng Y, Pan Q, Wan R. RNA demethylase ALKBH5 prevents pancreatic cancer progression by posttranscriptional activation of PER1 in an m6A-YTHDF2-dependent manner. Mol Cancer. 2020; 19:91. https://doi.org/10.1186/s12943-020-01158-w [PubMed]
- 124. Sun R, Yuan L, Jiang Y, Wan Y, Ma X, Yang J, Sun G, Zhou S, Wang H, Qiu J, Zhang L, Cheng W. ALKBH5 activates FAK signaling through m6A demethylation in ITGB1 mRNA and enhances tumor-associated lymphangiogenesis and lymph node metastasis in ovarian cancer. Theranostics. 2023; 13:833–48. https://doi.org/10.7150/thno.77441 [PubMed]
- 125. Li F, Zhang Y, Peng Z, Wang Y, Zeng Z, Tang Z. Diagnostic, clustering, and immune cell infiltration analysis of m6A regulators in patients with sepsis. Sci Rep. 2023; 13:2532. https://doi.org/10.1038/s41598-022-27039-4 [PubMed]
- 126. Lu N, Li X, Yu J, Li Y, Wang C, Zhang L, Wang T, Zhong X. Curcumin Attenuates Lipopolysaccharide-Induced Hepatic Lipid Metabolism Disorder by Modification of m6 A RNA Methylation in Piglets. Lipids. 2018; 53:53–63. https://doi.org/10.1002/lipd.12023 [PubMed]
- 127. Han YC, Xie HZ, Lu B, Xiang RL, Zhang HP, Li JY, Zhang SY. Lipopolysaccharide Alters the m6A Epitranscriptomic Tagging of RNAs in Cardiac Tissue. Front Mol Biosci. 2021; 8:670160. https://doi.org/10.3389/fmolb.2021.670160 [PubMed]
- 128. Liu M, Xu K, Saaoud F, Shao Y, Zhang R, Lu Y, Sun Y, Drummer C
4th , Li L, Wu S, Kunapuli SP, Criner GJ, Sun J, et al. 29 m6A-RNA Methylation (Epitranscriptomic) Regulators Are Regulated in 41 Diseases including Atherosclerosis and Tumors Potentially via ROS Regulation - 102 Transcriptomic Dataset Analyses. J Immunol Res. 2022; 2022:1433323. https://doi.org/10.1155/2022/1433323 [PubMed] - 129. Shen H, Xie K, Li M, Yang Q, Wang X. N6-methyladenosine (m6A) methyltransferase METTL3 regulates sepsis-induced myocardial injury through IGF2BP1/HDAC4 dependent manner. Cell Death Discov. 2022; 8:322. https://doi.org/10.1038/s41420-022-01099-x [PubMed]
- 130. Feng Z, Li Q, Meng R, Yi B, Xu Q. METTL3 regulates alternative splicing of MyD88 upon the lipopolysaccharide-induced inflammatory response in human dental pulp cells. J Cell Mol Med. 2018; 22:2558–68. https://doi.org/10.1111/jcmm.13491 [PubMed]
- 131. Wang B, Liu Y, Jiang R, Liu Z, Gao H, Chen F, Mei J. Emodin relieves the inflammation and pyroptosis of lipopolysaccharide-treated 1321N1 cells by regulating methyltransferase-like 3 -mediated NLR family pyrin domain containing 3 expression. Bioengineered. 2022; 13:6740–9. https://doi.org/10.1080/21655979.2022.2045836 [PubMed]
- 132. Dubey PK, Patil M, Singh S, Dubey S, Ahuja P, Verma SK, Krishnamurthy P. Increased m6A-RNA methylation and FTO suppression is associated with myocardial inflammation and dysfunction during endotoxemia in mice. Mol Cell Biochem. 2022; 477:129–41. https://doi.org/10.1007/s11010-021-04267-2 [PubMed]
- 133. Luo S, Liao C, Zhang L, Ling C, Zhang X, Xie P, Su G, Chen Z, Zhang L, Lai T, Tang J. METTL3-mediated m6A mRNA methylation regulates neutrophil activation through targeting TLR4 signaling. Cell Rep. 2023; 42:112259. https://doi.org/10.1016/j.celrep.2023.112259 [PubMed]
- 134. Shen H, Xie K, Tian Y, Wang X. N6-methyladenosine writer METTL3 accelerates the sepsis-induced myocardial injury by regulating m6A-dependent ferroptosis. Apoptosis. 2023; 28:514–24. https://doi.org/10.1007/s10495-022-01808-y [PubMed]
- 135. Zhang H, Liu J, Zhou Y, Qu M, Wang Y, Guo K, Shen R, Sun Z, Cata JP, Yang S, Chen W, Miao C. Neutrophil extracellular traps mediate m6A modification and regulates sepsis-associated acute lung injury by activating ferroptosis in alveolar epithelial cells. Int J Biol Sci. 2022; 18:3337–57. https://doi.org/10.7150/ijbs.69141 [PubMed]
- 136. Chen Y, Wu Y, Zhu L, Chen C, Xu S, Tang D, Jiao Y, Yu W. METTL3-Mediated N6-Methyladenosine Modification of Trim59 mRNA Protects Against Sepsis-Induced Acute Respiratory Distress Syndrome. Front Immunol. 2022; 13:897487. https://doi.org/10.3389/fimmu.2022.897487 [PubMed]
- 137. Luo J, Wang F, Sun F, Yue T, Zhou Q, Yang C, Rong S, Yang P, Xiong F, Yu Q, Zhang S, Wang CY, Li J. Targeted Inhibition of FTO Demethylase Protects Mice Against LPS-Induced Septic Shock by Suppressing NLRP3 Inflammasome. Front Immunol. 2021; 12:663295. https://doi.org/10.3389/fimmu.2021.663295 [PubMed]
- 138. Du J, Liao W, Liu W, Deb DK, He L, Hsu PJ, Nguyen T, Zhang L, Bissonnette M, He C, Li YC. N6-Adenosine Methylation of Socs1 mRNA Is Required to Sustain the Negative Feedback Control of Macrophage Activation. Dev Cell. 2020; 55:737–53.e7. https://doi.org/10.1016/j.devcel.2020.10.023 [PubMed]
- 139. Cheng L, Li H, Zhan H, Liu Y, Li X, Huang Y, Wang L, Zhang F, Li Y. Alterations of m6A RNA methylation regulators contribute to autophagy and immune infiltration in primary Sjögren’s syndrome. Front Immunol. 2022; 13:949206. https://doi.org/10.3389/fimmu.2022.949206 [PubMed]
- 140. Li G, Song Y, Liao Z, Wang K, Luo R, Lu S, Zhao K, Feng X, Liang H, Ma L, Wang B, Ke W, Yin H, et al. Bone-derived mesenchymal stem cells alleviate compression-induced apoptosis of nucleus pulposus cells by N6 methyladenosine of autophagy. Cell Death Dis. 2020; 11:103. https://doi.org/10.1038/s41419-020-2284-8 [PubMed]
- 141. Jin S, Zhang X, Miao Y, Liang P, Zhu K, She Y, Wu Y, Liu DA, Huang J, Ren J, Cui J. m6A RNA modification controls autophagy through upregulating ULK1 protein abundance. Cell Res. 2018; 28:955–7. https://doi.org/10.1038/s41422-018-0069-8 [PubMed]
- 142. Chao X, Wang S, Zhao K, Li Y, Williams JA, Li T, Chavan H, Krishnamurthy P, He XC, Li L, Ballabio A, Ni HM, Ding WX. Impaired TFEB-Mediated Lysosome Biogenesis and Autophagy Promote Chronic Ethanol-Induced Liver Injury and Steatosis in Mice. Gastroenterology. 2018; 155:865–79.e12. https://doi.org/10.1053/j.gastro.2018.05.027 [PubMed]
- 143. Settembre C, Di Malta C, Polito VA, Garcia Arencibia M, Vetrini F, Erdin S, Erdin SU, Huynh T, Medina D, Colella P, Sardiello M, Rubinsztein DC, Ballabio A. TFEB links autophagy to lysosomal biogenesis. Science. 2011; 332:1429–33. https://doi.org/10.1126/science.1204592 [PubMed]
- 144. Song H, Feng X, Zhang H, Luo Y, Huang J, Lin M, Jin J, Ding X, Wu S, Huang H, Yu T, Zhang M, Hong H, et al. METTL3 and ALKBH5 oppositely regulate m6A modification of TFEB mRNA, which dictates the fate of hypoxia/reoxygenation-treated cardiomyocytes. Autophagy. 2019; 15:1419–37. https://doi.org/10.1080/15548627.2019.1586246 [PubMed]
- 145. Wu T, Shao Y, Li X, Wu T, Yu L, Liang J, Zhang Y, Wang J, Sun T, Zhu Y, Chang X, Wang S, Chen F, Han X. NR3C1/Glucocorticoid receptor activation promotes pancreatic β-cell autophagy overload in response to glucolipotoxicity. Autophagy. 2023; 19:2538–57. https://doi.org/10.1080/15548627.2023.2200625 [PubMed]
- 146. Wang X, Wu R, Liu Y, Zhao Y, Bi Z, Yao Y, Liu Q, Shi H, Wang F, Wang Y. m6A mRNA methylation controls autophagy and adipogenesis by targeting Atg5 and Atg7. Autophagy. 2020; 16:1221–35. https://doi.org/10.1080/15548627.2019.1659617 [PubMed]
- 147. Cui YH, Yang S, Wei J, Shea CR, Zhong W, Wang F, Shah P, Kibriya MG, Cui X, Ahsan H, He C, He YY. Autophagy of the m6A mRNA demethylase FTO is impaired by low-level arsenic exposure to promote tumorigenesis. Nat Commun. 2021; 12:2183. https://doi.org/10.1038/s41467-021-22469-6 [PubMed]
- 148. Chen X, Gong W, Shao X, Shi T, Zhang L, Dong J, Shi Y, Shen S, Qin J, Jiang Q, Guo B. METTL3-mediated m6A modification of ATG7 regulates autophagy-GATA4 axis to promote cellular senescence and osteoarthritis progression. Ann Rheum Dis. 2022; 81:87–99. https://doi.org/10.1136/annrheumdis-2021-221091 [PubMed]
- 149. Li Q, Ni Y, Zhang L, Jiang R, Xu J, Yang H, Hu Y, Qiu J, Pu L, Tang J, Wang X. HIF-1α-induced expression of m6A reader YTHDF1 drives hypoxia-induced autophagy and malignancy of hepatocellular carcinoma by promoting ATG2A and ATG14 translation. Signal Transduct Target Ther. 2021; 6:76. https://doi.org/10.1038/s41392-020-00453-8 [PubMed]
- 150. Huang Y, Gao D, Wu Y, Sun L, Chen J, Chen J, Huang X, Yang J, Li S. YTHDF1 Protects Auditory Hair Cells from Cisplatin-Induced Damage by Activating Autophagy via the Promotion of ATG14 Translation. Mol Neurobiol. 2022; 59:7134–51. https://doi.org/10.1007/s12035-022-03021-z [PubMed]
- 151. Shen M, Li Y, Wang Y, Shao J, Zhang F, Yin G, Chen A, Zhang Z, Zheng S. N6-methyladenosine modification regulates ferroptosis through autophagy signaling pathway in hepatic stellate cells. Redox Biol. 2021; 47:102151. https://doi.org/10.1016/j.redox.2021.102151 [PubMed]
- 152. He M, Lei H, He X, Liu Y, Wang A, Ren Z, Liu X, Yan G, Wang W, Wang Y, Li G, Wang T, Pu J, et al. METTL14 Regulates Osteogenesis of Bone Marrow Mesenchymal Stem Cells via Inducing Autophagy Through m6A/IGF2BPs/Beclin-1 Signal Axis. Stem Cells Transl Med. 2022; 11:987–1001. https://doi.org/10.1093/stcltm/szac049 [PubMed]
- 153. Zheng C, Yu G, Su Q, Wu L, Tang J, Lin X, Chen Y, Guo Z, Zheng F, Zheng H, Lin L, Tang Y, Wu S, Li H. The deficiency of N6-methyladenosine demethylase ALKBH5 enhances the neurodegenerative damage induced by cobalt. Sci Total Environ. 2023; 881:163429. https://doi.org/10.1016/j.scitotenv.2023.163429 [PubMed]
- 154. Kageyama S, Gudmundsson SR, Sou YS, Ichimura Y, Tamura N, Kazuno S, Ueno T, Miura Y, Noshiro D, Abe M, Mizushima T, Miura N, Okuda S, et al. p62/SQSTM1-droplet serves as a platform for autophagosome formation and anti-oxidative stress response. Nat Commun. 2021; 12:16. https://doi.org/10.1038/s41467-020-20185-1 [PubMed]
- 155. Wu B, Song H, Fan M, You F, Zhang L, Luo J, Li J, Wang L, Li C, Yuan M. Luteolin attenuates sepsis-induced myocardial injury by enhancing autophagy in mice. Int J Mol Med. 2020; 45:1477–87. https://doi.org/10.3892/ijmm.2020.4536 [PubMed]
- 156. Chen Y, Wang J, Xu D, Xiang Z, Ding J, Yang X, Li D, Han X. m6A mRNA methylation regulates testosterone synthesis through modulating autophagy in Leydig cells. Autophagy. 2021; 17:457–75. https://doi.org/10.1080/15548627.2020.1720431 [PubMed]
- 157. Li G, Deng L, Huang N, Cui Z, Wu Q, Ma J, Pan Q, Sun F. m6A mRNA Methylation Regulates LKB1 to Promote Autophagy of Hepatoblastoma Cells through Upregulated Phosphorylation of AMPK. Genes (Basel). 2021; 12:1747. https://doi.org/10.3390/genes12111747 [PubMed]
- 158. Lu Z, Liu H, Song N, Liang Y, Zhu J, Chen J, Ning Y, Hu J, Fang Y, Teng J, Zou J, Dai Y, Ding X. METTL14 aggravates podocyte injury and glomerulopathy progression through N6-methyladenosine-dependent downregulating of Sirt1. Cell Death Dis. 2021; 12:881. https://doi.org/10.1038/s41419-021-04156-y [PubMed]
- 159. Huang J, Sun M, Tao Y, Ren J, Peng M, Jing Y, Xiao Q, Yang J, Lin C, Lei L, Yang Z, Zhang L. Cytoplasmic Expression of TP53INP2 Modulated by Demethylase FTO and Mutant NPM1 Promotes Autophagy in Leukemia Cells. Int J Mol Sci. 2023; 24:1624. https://doi.org/10.3390/ijms24021624 [PubMed]
- 160. Salcher S, Hermann M, Kiechl-Kohlendorfer U, Ausserlechner MJ, Obexer P. C10ORF10/DEPP-mediated ROS accumulation is a critical modulator of FOXO3-induced autophagy. Mol Cancer. 2017; 16:95. https://doi.org/10.1186/s12943-017-0661-4 [PubMed]
- 161. Lin Z, Niu Y, Wan A, Chen D, Liang H, Chen X, Sun L, Zhan S, Chen L, Cheng C, Zhang X, Bu X, He W, Wan G. RNA m6 A methylation regulates sorafenib resistance in liver cancer through FOXO3-mediated autophagy. EMBO J. 2020; 39:e103181. https://doi.org/10.15252/embj.2019103181 [PubMed]
- 162. Peng Z, Gong Y, Wang X, He W, Wu L, Zhang L, Xiong L, Huang Y, Su L, Shi P, Cao X, Liu R, Li Y, Xiao H. METTL3-m6A-Rubicon axis inhibits autophagy in nonalcoholic fatty liver disease. Mol Ther. 2022; 30:932–46. https://doi.org/10.1016/j.ymthe.2021.09.016 [PubMed]
- 163. Wu D, Wang H, Teng T, Duan S, Ji A, Li Y. Hydrogen sulfide and autophagy: A double edged sword. Pharmacol Res. 2018; 131:120–7. https://doi.org/10.1016/j.phrs.2018.03.002 [PubMed]
- 164. Yang L, Yan B, Qu L, Ren J, Li Q, Wang J, Kan X, Liu M, Wang Y, Sun Y, Wang C, Wang P. IGF2BP3 Regulates TMA7-mediated Autophagy and Cisplatin Resistance in Laryngeal Cancer via m6A RNA Methylation. Int J Biol Sci. 2023; 19:1382–400. https://doi.org/10.7150/ijbs.80921 [PubMed]
- 165. Han S, Zhu L, Zhu Y, Meng Y, Li J, Song P, Yousafzai NA, Feng L, Chen M, Wang Y, Jin H, Wang X. Targeting ATF4-dependent pro-survival autophagy to synergize glutaminolysis inhibition. Theranostics. 2021; 11:8464–79. https://doi.org/10.7150/thno.60028 [PubMed]
- 166. Li Y, Lu R, Niu Z, Wang D, Wang X. Suxiao Jiuxin Pill alleviates myocardial ischemia-reperfusion injury through the ALKBH5/GSK3β/mTOR pathway. Chin Med. 2023; 18:31. https://doi.org/10.1186/s13020-023-00736-6 [PubMed]
- 167. Li Z, Song Y, Liu L, Hou N, An X, Zhan D, Li Y, Zhou L, Li P, Yu L, Xia J, Zhang Y, Wang J, Yang X. miR-199a impairs autophagy and induces cardiac hypertrophy through mTOR activation. Cell Death Differ. 2017; 24:1205–13. https://doi.org/10.1038/cdd.2015.95 [PubMed]
- 168. Bai X, Wu J, Zhang M, Xu Y, Duan L, Yao K, Zhang J, Bo J, Zhao Y, Xu G, Zu H. DHCR24 Knock-Down Induced Tau Hyperphosphorylation at Thr181, Ser199, Thr231, Ser262, Ser396 Epitopes and Inhibition of Autophagy by Overactivation of GSK3β/mTOR Signaling. Front Aging Neurosci. 2021; 13:513605. https://doi.org/10.3389/fnagi.2021.513605 [PubMed]
- 169. Gao Z, Li C, Sun H, Bian Y, Cui Z, Wang N, Wang Z, Yang Y, Liu Z, He Z, Li B, Li F, Li Z, Wang L, Zhang D, Yang L, Xu Z, Li X, Xu H. N6-methyladenosine-modified USP13 induces pro-survival autophagy and imatinib resistance via regulating the stabilization of autophagy-related protein 5 in gastrointestinal stromal tumors. Cell Death Differ. 2023; 30:544–59. https://doi.org/10.1038/s41418-022-01107-8 [PubMed]
- 170. Sun Y, Shen W, Hu S, Lyu Q, Wang Q, Wei T, Zhu W, Zhang J. METTL3 promotes chemoresistance in small cell lung cancer by inducing mitophagy. J Exp Clin Cancer Res. 2023; 42:65. https://doi.org/10.1186/s13046-023-02638-9 [PubMed]
- 171. Mizushima N, Levine B. Autophagy in mammalian development and differentiation. Nat Cell Biol. 2010; 12:823–30. https://doi.org/10.1038/ncb0910-823 [PubMed]
- 172. David L, Taieb F, Pénary M, Bordignon PJ, Planès R, Bagayoko S, Duplan-Eche V, Meunier E, Oswald E. Outer membrane vesicles produced by pathogenic strains of Escherichia coli block autophagic flux and exacerbate inflammasome activation. Autophagy. 2022; 18:2913–25. https://doi.org/10.1080/15548627.2022.2054040 [PubMed]
- 173. Li Y, Xu J, Chen W, Wang X, Zhao Z, Li Y, Zhang L, Jiao J, Yang Q, Ding Q, Yang P, Wei L, Chen Y, et al. Hepatocyte CD36 modulates UBQLN1-mediated proteasomal degradation of autophagic SNARE proteins contributing to septic liver injury. Autophagy. 2023; 19:2504–19. https://doi.org/10.1080/15548627.2023.2196876 [PubMed]
- 174. Kang XF, Lu XL, Bi CF, Hu XD, Li Y, Li JK, Yang LS, Liu J, Ma L, Zhang JF. Xuebijing injection protects sepsis induced myocardial injury by mediating TLR4/NF-κB/IKKα and JAK2/STAT3 signaling pathways. Aging (Albany NY). 2023; 15:8501–17. https://doi.org/10.18632/aging.204990 [PubMed]
- 175. Zhao H, Chen H, Xiaoyin M, Yang G, Hu Y, Xie K, Yu Y. Autophagy Activation Improves Lung Injury and Inflammation in Sepsis. Inflammation. 2019; 42:426–39. https://doi.org/10.1007/s10753-018-00952-5 [PubMed]
- 176. Zhu DD, Huang YL, Guo SY, Li N, Yang XW, Sui AR, Wu Q, Zhang Y, Kong Y, Li QF, Zhang T, Zheng WF, Li AP, Yu J, Ma TH, Li S. AQP4 Aggravates Cognitive Impairment in Sepsis-Associated Encephalopathy through Inhibiting Nav1.6-Mediated Astrocyte Autophagy. Adv Sci (Weinh). 2023; 10:e2306241. https://doi.org/10.1002/advs.202306241 [PubMed]
- 177. Martino E, D’Onofrio N, Balestrieri A, Mele L, Sardu C, Marfella R, Campanile G, Balestrieri ML. MiR-15b-5p and PCSK9 inhibition reduces lipopolysaccharide-induced endothelial dysfunction by targeting SIRT4. Cell Mol Biol Lett. 2023; 28:66. https://doi.org/10.1186/s11658-023-00482-5 [PubMed]
- 178. Zhang W, Chen H, Xu Z, Zhang X, Tan X, He N, Shen J, Dong J. Liensinine pretreatment reduces inflammation, oxidative stress, apoptosis, and autophagy to alleviate sepsis acute kidney injury. Int Immunopharmacol. 2023; 122:110563. https://doi.org/10.1016/j.intimp.2023.110563 [PubMed]
- 179. Fan H, Cui J, Liu F, Zhang W, Yang H, He N, Dong Z, Dong J. Malvidin protects against lipopolysaccharide-induced acute liver injury in mice via regulating Nrf2 and NLRP3 pathways and suppressing apoptosis and autophagy. Eur J Pharmacol. 2022; 933:175252. https://doi.org/10.1016/j.ejphar.2022.175252 [PubMed]
- 180. Kumar V. Sepsis roadmap: What we know, what we learned, and where we are going. Clin Immunol. 2020; 210:108264. https://doi.org/10.1016/j.clim.2019.108264 [PubMed]
- 181. Bi CF, Liu J, Hao SW, Xu ZX, Ma X, Kang XF, Yang LS, Zhang JF. Xuebijing injection protects against sepsis-induced myocardial injury by regulating apoptosis and autophagy via mediation of PI3K/AKT/mTOR signaling pathway in rats. Aging (Albany NY). 2023; 15:4374–90. https://doi.org/10.18632/aging.204740 [PubMed]
- 182. Zaccara S, Ries RJ, Jaffrey SR. Reading, writing and erasing mRNA methylation. Nat Rev Mol Cell Biol. 2019; 20:608–24. https://doi.org/10.1038/s41580-019-0168-5 [PubMed]
- 183. Chen X, Wang J, Tahir M, Zhang F, Ran Y, Liu Z, Wang J. Current insights into the implications of m6A RNA methylation and autophagy interaction in human diseases. Cell Biosci. 2021; 11:147. https://doi.org/10.1186/s13578-021-00661-x [PubMed]
- 184. Zhang H, Wu D, Wang Y, Guo K, Spencer CB, Ortoga L, Qu M, Shi Y, Shao Y, Wang Z, Cata JP, Miao C. METTL3-mediated N6-methyladenosine exacerbates ferroptosis via m6A-IGF2BP2-dependent mitochondrial metabolic reprogramming in sepsis-induced acute lung injury. Clin Transl Med. 2023; 13:e1389. https://doi.org/10.1002/ctm2.1389 [PubMed]
- 185. Song B, Zeng Y, Cao Y, Zhang J, Xu C, Pan Y, Zhao X, Liu J. Emerging role of METTL3 in inflammatory diseases: mechanisms and therapeutic applications. Front Immunol. 2023; 14:1221609. https://doi.org/10.3389/fimmu.2023.1221609 [PubMed]
- 186. Wang C, Meng Y, Zhao J, Ma J, Zhao Y, Gao R, Liu W, Zhou X. Deubiquitinase USP13 regulates glycolytic reprogramming and progression in osteosarcoma by stabilizing METTL3/m6A/ATG5 axis. Int J Biol Sci. 2023; 19:2289–303. https://doi.org/10.7150/ijbs.82081 [PubMed]
- 187. He Y, Wang W, Xu X, Yang B, Yu X, Wu Y, Wang J. Mettl3 inhibits the apoptosis and autophagy of chondrocytes in inflammation through mediating Bcl2 stability via Ythdf1-mediated m6A modification. Bone. 2022; 154:116182. https://doi.org/10.1016/j.bone.2021.116182 [PubMed]
- 188. Qu M, Chen Z, Qiu Z, Nan K, Wang Y, Shi Y, Shao Y, Zhong Z, Zhu S, Guo K, Chen W, Lu X, Wang Z, et al. Neutrophil extracellular traps-triggered impaired autophagic flux via METTL3 underlies sepsis-associated acute lung injury. Cell Death Discov. 2022; 8:375. https://doi.org/10.1038/s41420-022-01166-3 [PubMed]