



Introduction
Hepatocellular carcinoma (HCC) is the most common pathological type of liver cancer and the leading cause of death in patients with liver cancer [1, 2]. The incidence and mortality rates of liver cancer worldwide rank 6th and 3rd of all cancers, respectively, with approximately 960,000 new cases and 830,000 deaths per year [3]. The main risk factors of HCC are hepatitis B virus (HBV) infection, hepatitis C virus (HCV) infection, alcohol consumption, obesity, diabetes, non-alcoholic fatty liver disease, aflatoxins, etc. [4, 5]. Although significant progress has been made in treating HCC with surgery, chemotherapy, targeted therapy, immunotherapy, and interventional therapy, the early symptoms of HCC are atypical, and most patients are at advanced stages when diagnosed. At this time, the available treatments are limited, resulting in a poor prognosis with a 5-year survival rate of approximately 18.4% [6, 7]. Therefore, it is importance to explore more effective therapeutic targets and identify novel prognostic markers to improve HCC survival.
Protein post-translational modifications (PTMs) are processes that employ reversible state patterns to modulate protein function and alter physicochemical properties [8]. Studies have identified more than 400 post-translational modifications, with common modifications including ubiquitination, sumoylation, phosphorylation, acetylation, and methylation [9]. The ubiquitin-proteasome system (UPS) is a common post-translational modification pathway that is involved in the regulation of cell survival and differentiation [10]. Ubiquitination requires the participation of three enzymes, namely ubiquitin-activating enzyme (E1), ubiquitin-binding enzyme (E2), and ubiquitin-protein ligase (E3) [11]. UBE4B belongs to the U-box family of ubiquitin ligases, also known as UFD2a, which is a novel E3 [12]. Many studies have investigated the substrate function of UBE4B, and it has been confirmed that it is involved in the ubiquitinated degradation of important proteins such as EGFR, p53, and caspase3 [13, 14]. Additionally, UBE4B can be involved in the development of various cancers, such as renal cell carcinoma, breast cancer, and neural tube cell tumors, by regulating the ubiquitination of substrate proteins [15–17]. These results suggest that UBE4B may be a novel prognostic indicator in patients with multiple malignancies.
In this study, we investigated the UBE4B expression, prognosis, tumor immune microenvironment, and regulatory mechanisms of HCC. Next, we explored non-coding RNA regulation associated with UBE4B and constructed a critical UBE4B-miRNA-LncRNA regulatory axis. These findings will aid us to understand the relevant functions of UBE4B and provide new strategies for drug development and exploration of prognosis-related biomarkers.
Materials and Methods
Data acquisition
The expression of UBE4B was evaluated in 33 cancer and normal tissues using the The Cancer Genome Atlas (TCGA) (https://www.cancer.gov/tcga) and The Genotype-Tissue Expression (GTEx) (http://commonfund.nih.gov/GTEx/) databases. RNA sequencing data and clinical follow-up information of 33 patients with cancer were obtained from the TCGA cohort. All expression data were normalized using a log2 transformation.
TIMER database analysis
Tumor Immune Estimation Resource (TIMER) (https://cistrome.shinyapps.io/timer/) is a web server for analyzing the abundance of tumor infiltrates [18]. It was used to investigate the expression and abundance of immune infiltration in different cancers based on the TCGA cohort, with the threshold value as log2TPM.
GEPIA database analysis
Gene Expression Profiling Interactive Analysis (GEPIA) (http://gepia.cancer-pku.cn/) is a web tool for cancer and normal gene-expression profiling and interactive analysis using TCGA and GTEx cohorts [19]. Analysis of violin images of UBE4B in various pathological stages were performed using the Pathological Stage Map module in the GEPIA database.
Kaplan–Meier plotter analysis
The relationship between UBE4B expression and prognosis, including overall survival (OS) and recurrence-free survival (RFS), was examined using the Kaplan–Meier plotter (http://kmplot.com/analysis/) in different cancer types. Hazard ratios (HRs) and 95% confidence intervals (CIs) were calculated along with the log-rank p-value.
Competing endogenous RNAs (CeRNA) network construction
TarBase V.8 (https://dianalab.e-ce.uth.gr/html/diana/web/index.php?r=tarbasev8) was used to predict the miRNA targets that bind to UBE4B. The lncRNAs upstream of these miRNAs were identified using miRNet (https://www.mirnet.ca/miRNet/home.xhtml). StarBase (https://starbase.sysu.edu.cn/index.php) was used to predict miRNA and lncRNA expression levels and their prognostic values. Additionally, starBase was used to perform expression correlation analysis of hsa-miR-22-3p-UBE4B, lncRNA-hsa-miR-22-3p, and lncRNA-UBE4B in HCC.
TISIDB database analysis
TISIDB (http://cis.hku.hk/TISIDB/index.php) is an online platform for the interaction between tumors and the immune system that integrates multiple heterogeneous data types [20]. The TISIDB database was used investigated the relationship between UBE4B and 45 immunostimulants, 24 immunosuppressive agents, 41 chemokines, and 18 receptors in HCC.
Cell culture
The human liver (LO2) and human HCC (7721 and HepG2) cell lines were purchased from Cell Bank (Shanghai, China). All cell lines were cultured in Dulbecco’s modified Eagle’s medium (DMEM; cat no.SH30022.01; HyClone) supplemented with 10% fetal bovine serum (FBS; cat no.2127186; VivaCell) and 1% penicillin and streptomycin (cat no.15140122; Gibco) at 37° C in 5% CO2.
Quantitative real-time PCR (qRT-PCR)
Total RNA was extracted from the human liver (LO2) and human HCC (7721 and HepG2) cell lines using TRIzol reagent (cat no.12183-555; Invitrogen). Reverse transcription of cDNA to cDNA using PrimeScript™ RT Master Mix kit (cat no.RR036A; Takara). qRT-PCR analysis was performed using TB Green® Premix Ex Taq™ II (cat no.RR820A; Takara). The primers used were as follows: UBE4B forward:5′-CTACCTCCCCAATAGGTGCAT-3′ and UBE4B reverse:5′-GGCGAGCTGCTGAGAGAAC-3′; FDG5-AS1 forward:5′-GAAGGGCCGAAGAGCTCAAT-3′ and FDG5-AS1 reverse:5′-GGCTCGCAAAGTGTCTGTTG-3′; LINC00858 forward:5′-CCCAGCTCCTTACACACGTT-3′ and LINC00858 reverse:5′-TTCAGAGGCCTGCATCACTG-3′; SNHG16 forward:5′-CAGAATGCCATGGTTTCCCC-3′ and SNHG16 reverse:5′-TGGCAAGAGACTTCCTGAGG-3′; Has-miR-22-3P forward:5′-TCAGTGCATCACAGAACTTTGT-3′ and Has-miR-22-3P reverse:5′-TGGCAAGAGACTTCCTGAGG-3′; Actin forward:5′-GTGGCCGAGGACTTTGATTG-3′ and Actin reverse:5′-CCTGTAACAACGCATCTCATATT-3′. Relative quantification was performed using the 2−ΔΔCt method.
HPA database analysis
The Human Protein Atlas (HPA) (www.proteinatlas.org/) was used to detect the protein expression of UBE4B in hepatocellular carcinoma and corresponding normal tissues in the “tissue” and “pathology” modules. All images were confirmed by immunohistochemical experiments and specific patient information is listed.
Statistical analysis
All data analyses were automatically performed using the online database mentioned above. Log-rank p-value <0.05 or p-value <0.05 were considered statistically significant.
Availability of data and materials
All data generated or analyzed during this study are included in this article.
Results
Identification of UBE4B expression in pan-cancer
The expression levels of UBE4B in tumors and normal tissues were investigated. The results suggested that UBE4B mRNA was overexpressed in Cholangiocarcinoma (CHOL), Head and Neck Squamous Cell Carcinoma (HNSC), Liver Hepatocellular Carcinoma (LIHC), Lung Adenocarcinoma (LUAD), and Lung Squamous Cell Carcinoma (LUSC) using the TIMER database and downregulated in Breast Invasive Carcinoma (BRCA), Kidney Chromophobe (KICH), Kidney Renal Clear Cell Carcinoma (KIRC), Kidney Renal Papillary Cell Carcinoma (KIRP), Thyroid Carcinoma (THCA), and Uterine Corpus Endometrial Carcinoma (UCEC) (Figure 1A). Then, samples were integrated from the GTEx database and the expression levels of UBE4B in various cancers were compared. UBE4B mRNA was upregulated in BRCA, CHOL, Lymphoid Neoplasm Diffuse Large B-cell Lymphoma (DLBC), HNSC, KIRC, KIRP, LIHC, LUAD, LUSC, Ovarian Serous Cystadenocarcinoma (OV), Pancreatic Adenocarcinoma (PAAD), Prostate Adenocarcinoma (PRAD), Skin Cutaneous Melanoma (SKCM), Stomach Adenocarcinoma (STAD), and Testicular Germ Cell Tumors (TGCT) and downregulated in Adrenocortical Carcinoma (ACC), Bladder Urothelial Carcinoma (BLCA), Colon Adenocarcinoma (COAD), Glioblastoma Multiforme (GBM), KICH, Brain Lower Grade Glioma (LGG), Rectum Adenocarcinoma (READ), THCA, and Uterine Carcinosarcoma (UCS) (Figure 1B). In the CPTAC database, UBE4B protein expression was markedly increased in BRCA, HNSC, LIHC, UCEC, and LUAD, and decreased in COAD, GBM, and KIRC compared to normal tissues (Figure 1C).
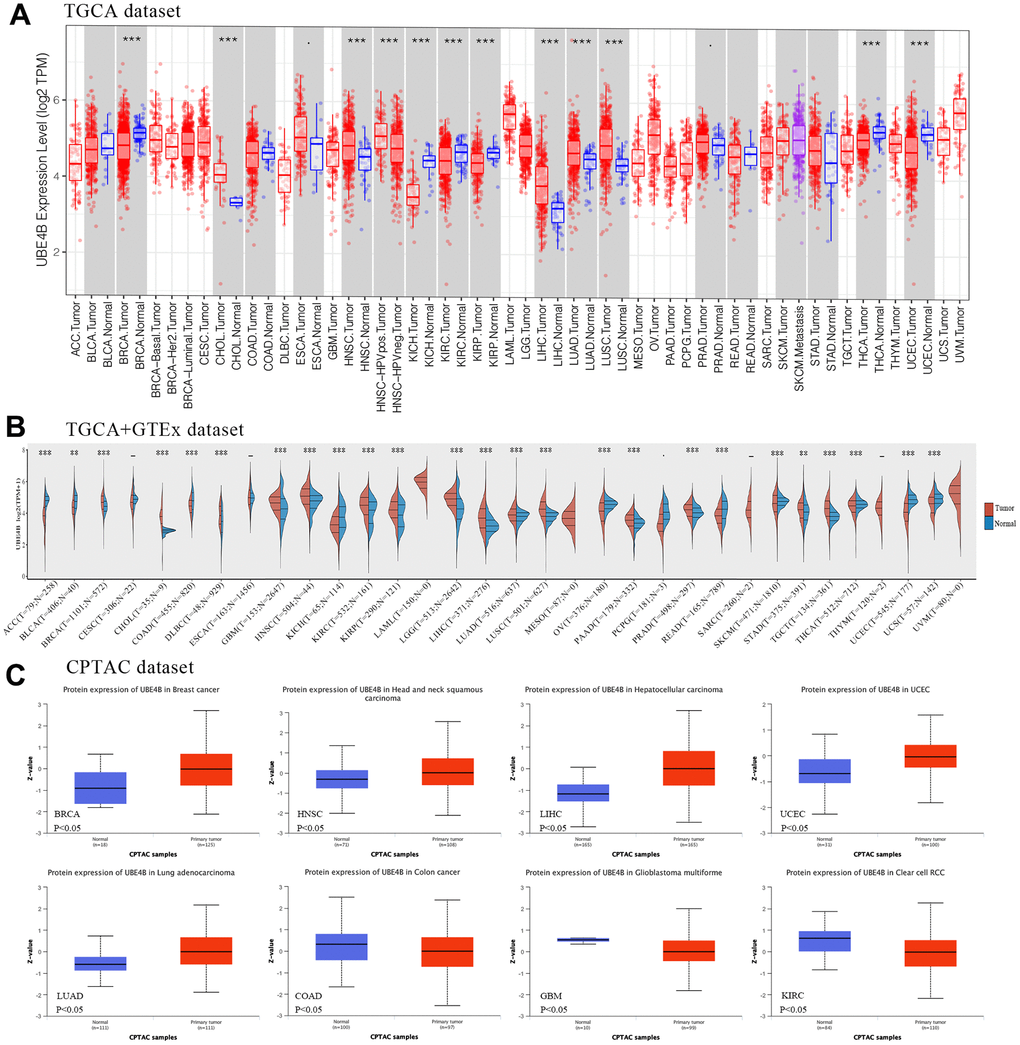
Figure 1. Expression of UBE4B in pan-cancer. (A) UBE4B expression in normal and cancer tissues in TIMER. (B) UBE4B expression in TCGA cancers compared with corresponding TCGA and GTEx normal tissues. (C) The UBE4B protein expression was determined by the CPTAC dataset in BRCA, HNSC, LIHC, UCEC, LUAD, COAD, GBM, and KIRC. *P < 0.05; **P < 0.01; ***P < 0.001.
Clinicopathology and prognostic value of UBE4B in pan-cancer
The GEPIA database was used to elucidate the relationship between UBE4B expression and pathological stages in different cancers, revealing significant correlations between UBE4B expression and pathological stages of COAD (P = 0.00308), KIRC (P = 8.01 × 10-5), LIHC (P = 0.00739), and OV (P = 0.0233) (Figure 2A). Next, pan-cancer survival analysis for UBE4B was performed. The OS results indicated that patients with BLCA (P = 0.001), LIHC (P = 7.4 × 10-5), and Sarcoma (SARC) (P = 0.00013) with high UBE4B expression exhibited a worse prognosis, while patients with Esophageal Carcinoma (ESCA) (P = 0.018), HNSC (P = 0.035), and KIRC (P = 6 × 10-6) exhibited an improved prognosis. Moreover, UBE4B overexpression was correlated with inferior prognosis in patients with LIHC (P = 0.03) and UCEC (P = 0.021) but with superior prognosis in patients with ESCA (P = 0.044) (Figure 2B).
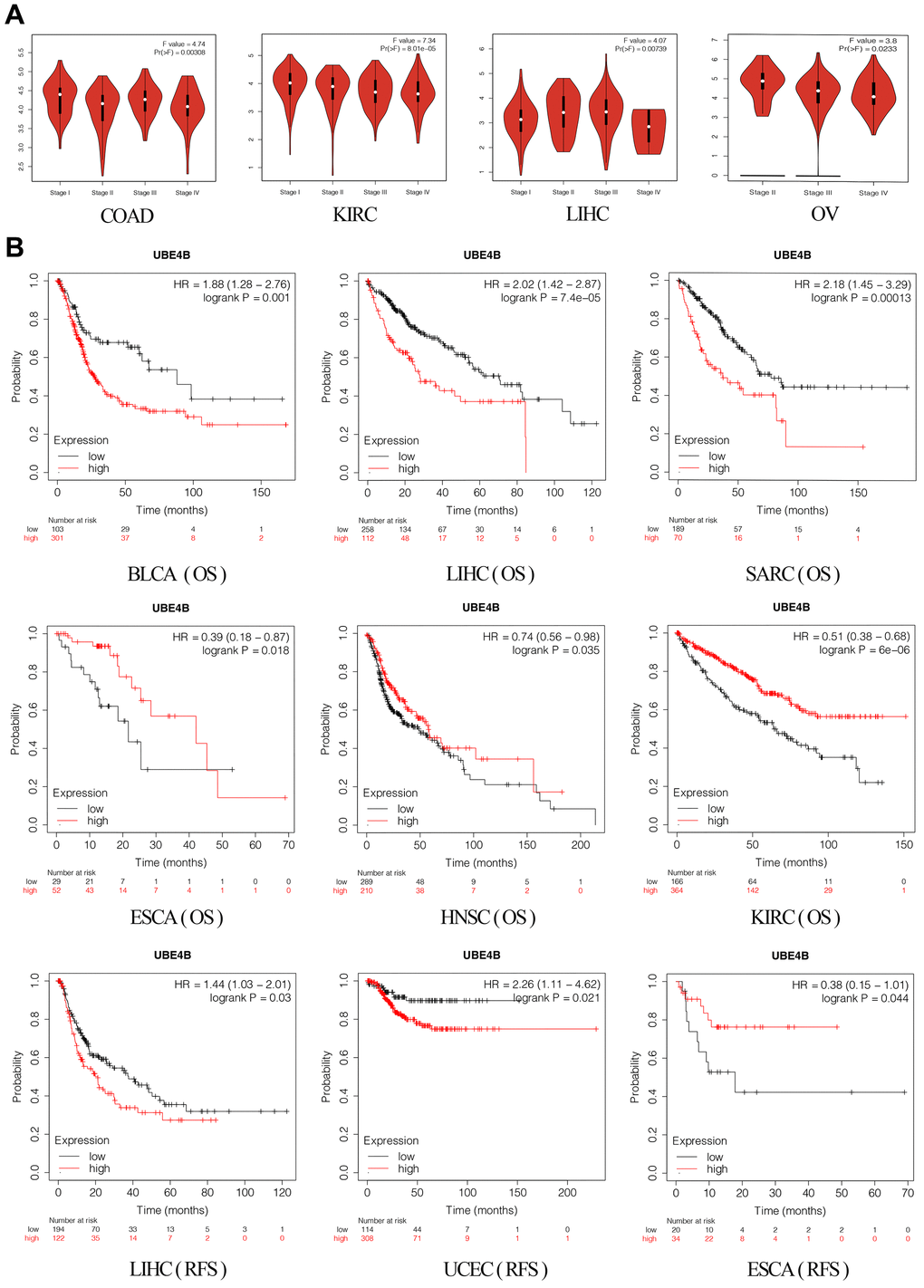
Figure 2. Correlation of UBE4B expression with pathological staging and prognosis. (A) Expression of UBE4B in pathological stages (stage I, stage II, stage III, and stage IV) of COAD, KIRC LIHC, and OV. (B) The OS and RFS plot of UBE4B in different cancers by Kaplan-Meier Plotter.
Forecast and evaluation of the upstream miRNAs of UBE4B
The miRNAs upstream of UBE4B was predicted using the TarBase V.8 database, and 55 miRNAs were retrieved (Figure 3A). In accordance with the ceRNA hypothesis, we estimated the predicted miRNA expression patterns and prognostic value in LIHC using the starBase database. The results indicated that only downregulation of hsa-miR-22-3p was correlated with worse OS in patients with LIHC (P = 5.4 × 10-10; P = 0.0013) (Figure 3B, 3C). A correlation analysis of UBE4B and hsa-miR-22-3p was conducted (Figure 3D), indicating that the hsa-miR-22-3p/UBE4B axis was involved in the ceRNA mechanism. Therefore, hsa-miR-22-3p could be considered as a key miRNA for follow-up exploration.
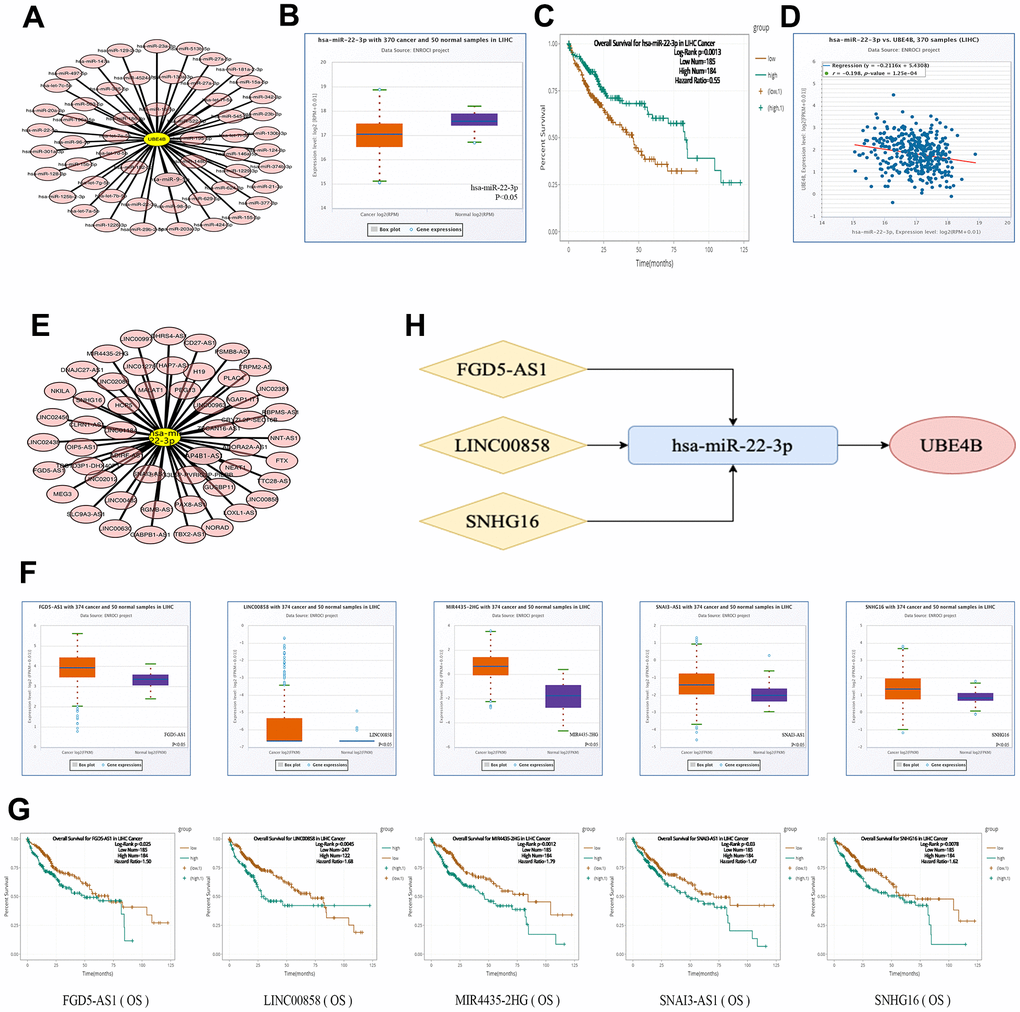
Figure 3. Construction of ceRNA network. (A) The miRNA-UBE4B regulatory network was developed by cytoscape software. (B–D) The expression (B), OS (C) and correlation with UBE4B (D) of hsa-miR-22-3p in HCC by StarBase database. (E) The lncRNA-hsa-miR-22-3p regulatory network constructed by cytoscape software. (F, G) The expression (F) and OS (G) of FGD5-AS1, LINC00858, MIR4435-2HG, SNAI3-AS1 and SNHG16 by StarBase database. (H) The schematic diagram of potential ceRNA network regulation axis.
Prediction and validation of upstream lncRNAs of hsa-miR-22-3p
The online miRNet database was used to predict the upstream lncRNA of hsa-miR-22-3p. A total of 52 lncRNAs were identified (Figure 3E). A ceRNA-based hypothesis argues that lncRNAs negatively regulate miRNAs, while positively modulating mRNA [21]. Moreover, we evaluated the expression and prognosis of the 52 lncRNAs using the starBase database. The expression levels of five lncRNAs (MIR4435-2HG, FGD5-AS1, LINC00858, SNAI3-AS1, and SNHG16) was significantly upregulated and associated with adverse prognosis in patients with LIHC (P < 0.05) (Figure 3F, 3G). Additionally, we analyzed the correlation between five lncRNAs and hsa-miR-22-3p or UBE4B using the starBase database and revealed that three lncRNAs (FGD5-AS1, LINC00858, and SNHG16) were inversely co-expressed with hsa-miR-22-3p and positively co-expressed with UBE4B (Table 1). We constructed a potential FGD5-AS1/LINC00858/SNHG16/hsa-miR-22-3p/UBE4B regulation axis, which may play a vital role in the development and progression of LIHC (Figure 3H).
Table 1. Correlation analysis between lncRNA and hsa-mir-22-3p or lncRNA and UBE4B in HCC determined by starBase database.
| lncRNA | miRNA | R value | p value | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MIR4435-2HG | hsa-mir-22-3p | -0.168 | 1.16E-03**a | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FGD5-AS1 | hsa-mir-22-3p | -0.304 | 2.30E-09***a | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LINC00858 | hsa-mir-22-3p | -0.196 | 1.47E-04***a | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| SNAI3-AS1 | hsa-mir-22-3p | -0.169 | 1.07E-03**a | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| SNHG16 | hsa-mir-22-3p | -0.382 | 2.62E-14***a | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| lncRNA | mRNA | R value | p value | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MIR4435-2HG | UBE4B | -0.046 | 3.74E-01 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FGD5-AS1 | UBE4B | 0.53 | 1.61E-28***a | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LINC00858 | UBE4B | 0.275 | 6.53E-08***a | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| SNAI3-AS1 | UBE4B | -0.003 | 9.47E-01 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| SNHG16 | UBE4B | 0.266 | 1.70E-07***a | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| aThese results are statistically significant. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| **P value < 0.01;***P value <0.001. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Correlation between immune infiltration and UBE4B expression in LIHC
Immune infiltration plays a pivotal role in tumor pathogenesis [22]. Immune cell infiltration did not change significantly at different UBE4B copy numbers in LIHC (Figure 4A). The association between UBE4B expression and the degree of immune cell infiltration was evaluated in LIHC using the TIMER database. A significant positive correlation was observed between UBE4B expression and immune cells in LIHC, including B cells (P = 3.21 × 10-2), CD8+ T cells (P = 1.59 × 10-2), CD4+ T cells (P = 9.30 × 10-6), macrophages (P = 9.19 × 10-7), neutrophils (P = 6.47 × 10-9), and dendritic cells (P = 4.97 × 10-7) (Figure 4B). Furthermore, the correlation between UBE4B expression and immune cell biomarkers was evaluated in patients with LIHC. A significant positive correlation was observed between UBE4B and most immune cell biomarkers in LIHC, in addition to CCR7, CD1C, CD79A, CD8A, CD8B, and HLA-DPB1 (Figure 4C).
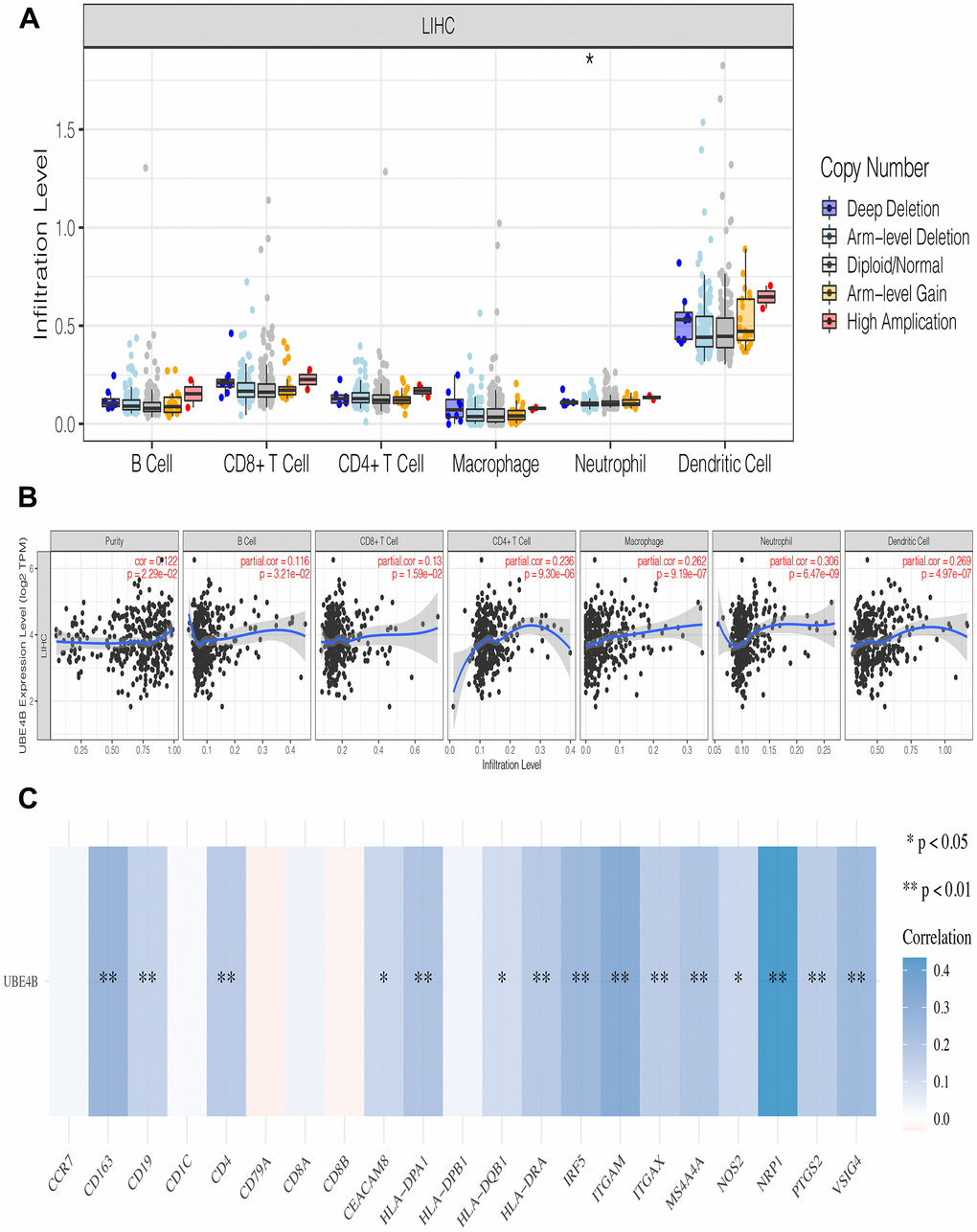
Figure 4. The relationship of immune cell infiltration with UBE4B level in HCC. (A) The infiltration level of various immune cells in HCC with different copy numbers of UBE4B. (B) The correlation of UBE4B expression level with B cell, CD8+ T cell, CD4+ T cell, macrophage, neutrophil, or dendritic cell infiltration level in HCC. (C) Correlation analysis between UBE4B and biomarkers of immune cells in HCC. *P < 0.05; **P < 0.01.
Relationship between UBE4B expression and immune modulators in LIHC
Immune modulators affect the function of the immune system [23]. UBE4B was significantly associated with most immune inhibitors, including ADORA2A, BTLA, CD96, CD160, CD244, CD274, CSF1R, CTLA4, HAVCR2, IL10RB, KDR, LAG3, LGALS9, PDCD1, PVRL2, TGFB1, TIGIT, and VTCN1 (P < 0.05) (Figure 5A). Moreover, UBE4B expression was closely associated with the following immune stimulators: C10orf54, CD27, CD40, CD40LG, CD48, CD86, CD276, CXCL12, CXCR4, ENTPD1, ICOS, ICOSLG, IL6, IL6R, KLRK1, LTA, NT5E, PVR, TMEM17, TNFRSF4, TNFRSF8, TNFRSF17, TNFRSF18, TNFRSF25, TNFSF13B, and TNFSF15 (P < 0.05) (Figure 5B). These results indicate the possible involvement of UBE4B in modulating tumor immune escape.
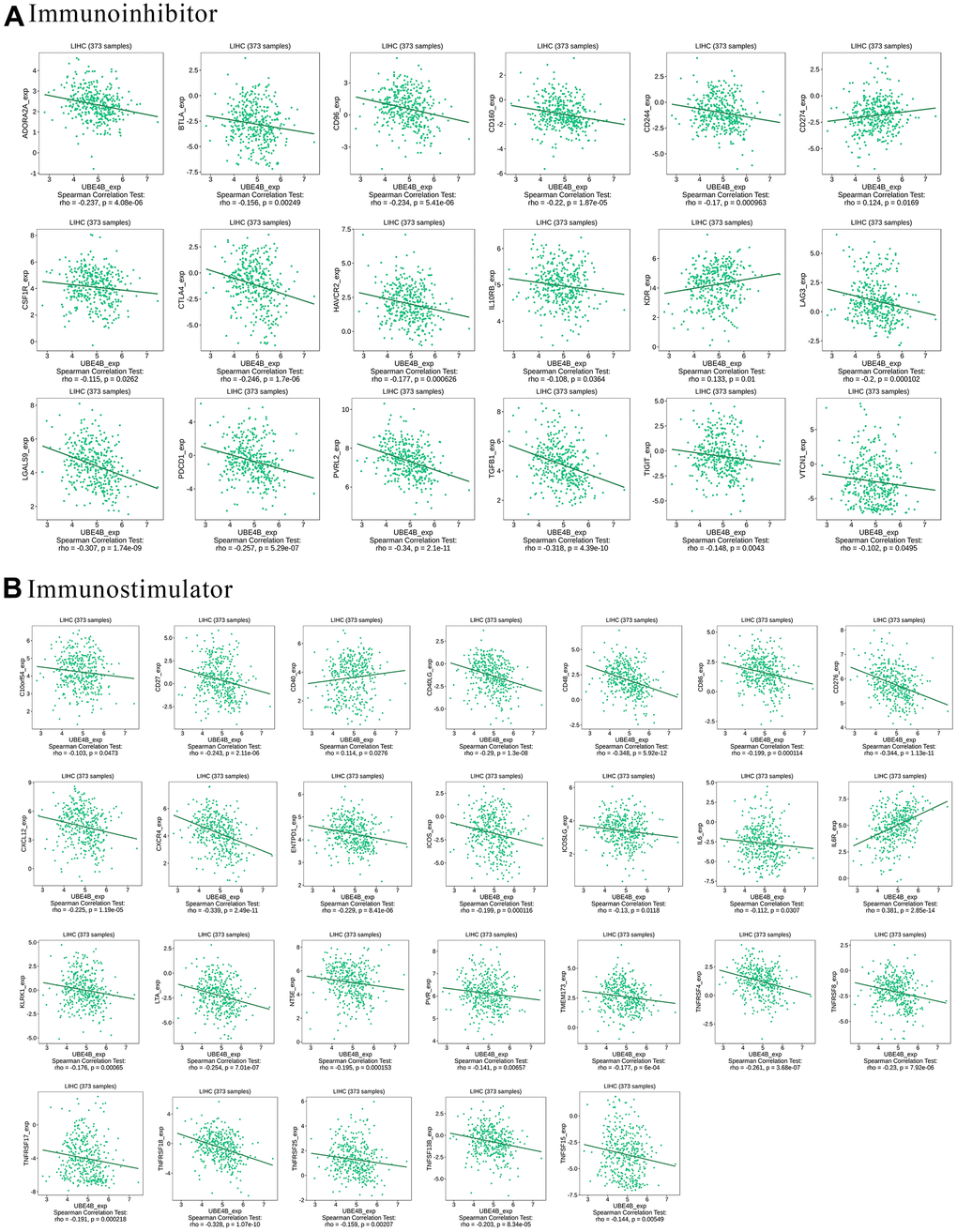
Figure 5. Correlation between UBE4B expression and immunomodulators in HCC by TISIDB database. (A) Immunoinhibitors, (B) immunostimulators.
Relationship between UBE4B expression and chemokines in LIHC
Chemokines enable directed chemotaxis of immune cells [24]. UBE4B expression was significantly associated with chemokines such as CCL2, CCL3, CCL4, CCL5, CCL11, CCL14, CCI18, CCL19, CCL20, CCL21, CCL22, CCL25, CCL26, CX3CL1, CXCL1, CXCL2, CXCL5, CXCL12, CXCL14, CXCL16, CXCL17, and XCL2 (P < 0.05) (Figure 6A). We further investigated the correlation between UBE4B and chemokine receptor expression. We found that UBE4B expression was significantly linked to CCR2, CCR5, CCR6, CCR7, CCR10, CX3CR1, CXCR1, CXCR3, CXCR4, CXCR5, and CXCR6 expression (P < 0.05) (Figure 6B). These results further demonstrated that UBE4B is involved in immune modulation in LIHC.
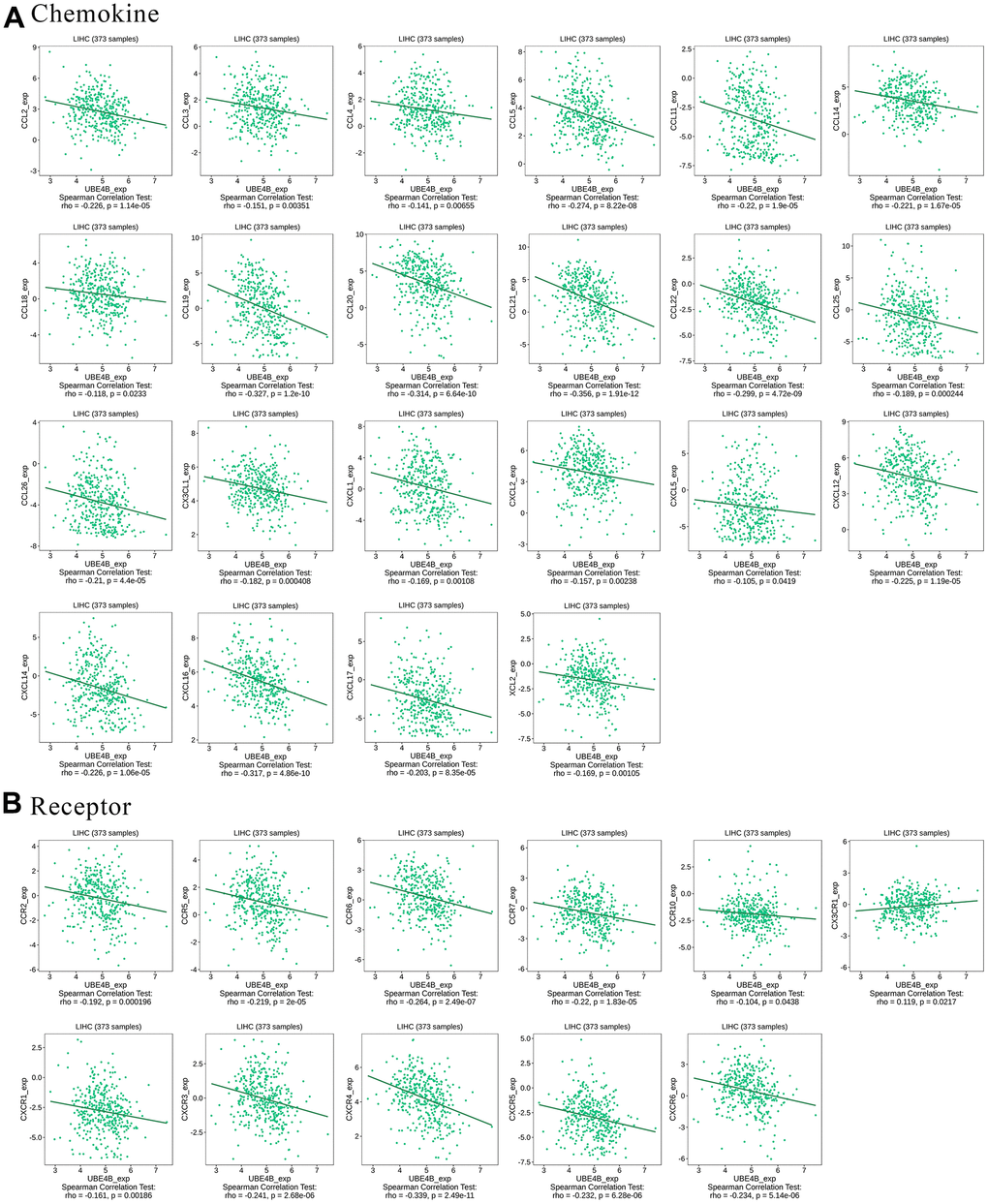
Figure 6. Correlation between UBE4B expression and chemokines in HCC by TISIDB database. (A) Chemokines, (B) chemokine receptors.
Verification of the mRNA and protein expression of genes
The mRNA expression levels of FGD5-AS1, LINC00858, SNHG16, hsa-miR-22-3p, and UBE4B were evaluated in HCC cell lines. The mRNA expression levels of FGD5-AS1, LINC00858, SNHG16, and UBE4B were elevated in the HCC cell lines (7721 and HepG2) compared to the normal liver cell line (LO2), while the mRNA expression of hsa-miR-22-3p showed a decreasing trend (Figure 7A–7E). Furthermore, we explored the protein expression of UBE4B in HCC using the HPA database. The results revealed that UBE4B protein expression was higher in HCC tissues than in corresponding normal tissues, mainly in the cytoplasm and cellular membranes (Figure 7F–7I).
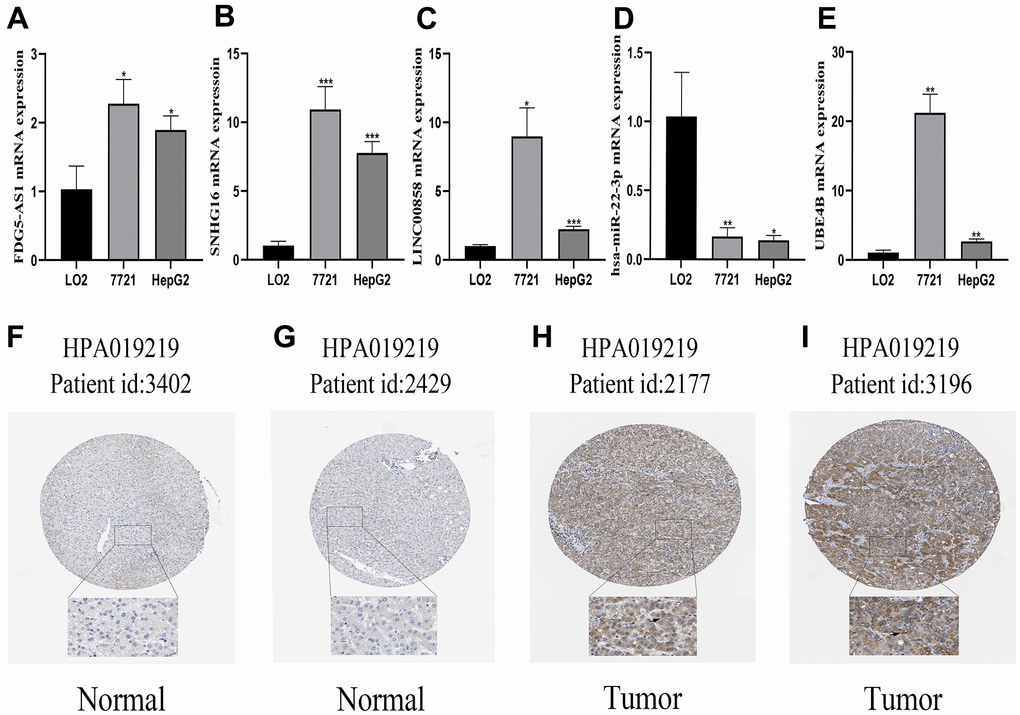
Figure 7. Verification of the mRNA and protein expression of genes. (A–E) qRT-PCR analysis detected the expression of FGD5-AS1, SNHG16, LINC00858, hsa-miR-22-3p, and UBE4B in normal cell line (LO2), and HCC cell lines (7721, HepG2). (F–I) Verification of the protein expression of UBE4B in HCC and corresponding normal tissues by HPA database. *P < 0.05; **P < 0.01; ***P < 0.001.
Discussion
HCC has been a problem affecting human health for many years. There are observable variations in tumor proliferation, invasion, and metastasis between different HCC subtypes; therefore, it is necessary to further investigate biological molecular regulation in HCC. UBE4B is a U-box family of E4-active ubiquitin ligases that determine the specific recognition of substrates [16]. UBE4B can act as an oncogene or tumor suppressor gene in different types of cancers with opposite functions [13, 25]. Zhang et al. showed that UBE4B influenced the development of HCC by promoting the growth of HCC cells, and silencing of UBE4B inhibited the proliferation, migration, and invasion of HCC cells, resulting in significant apoptosis [26]. However, a regulatory axis related to UBE4B in HCC is still lacking and requires detailed exploration.
In this study, the expression of UBE4B was analyzed in pan-cancer using TCGA and GTEx databases. UBE4B expression was found to be increased at both mRNA and protein levels in HCC compared with normal samples, and was associated with clinicopathological characteristics in the survival analysis. OS analysis also indicated the prognostic value of UBE4B in several cancers, including BLCA, LIHC, SARC, ESCA, HNSC, and KIRC. A previous study demonstrated that UBE4B expression is often upregulated at the transcriptional (71%) and translational (84%) levels in HCC tissues [16]. This report was consistent with our results, which illustrate the carcinogenic effects of UBE4B in HCC.
The correlation between UBE4B expression and HCC progression was investigated. UBE4B upstream miRNA expression patterns, more specifically hsa-miR-22-3p, also affected HCC in the starBase database. Recent studies have found that hsa-miR-22-3p acts as an important regulatory gene in breast cancer [27], Japanese encephalitis [28], and obesity [29]. The expression and prognosis of the upstream lncRNA of hsa-miR-22-3p, as a negative precursor molecule for miRNA, was analyzed using the starBase database, and five related lncRNAs (MIR4435-2HG, FGD5-AS1, LINC00858, SNAI3-AS1, and SNHG16) were shown to be significantly upregulated in HCC and associated with poor prognosis in patients with HCC. This is a top-down, molecular regulatory mechanism.
P53 protein, a well-known substrate of UBE4B, has been shown to regulate cell cycle arrest, apoptosis, and DNA repair processes, which play critical roles in preventing tumor progression [30, 31]. This indicates that the overexpression of UBE4B leads to a carcinogenic process, with the amount of p53 protein greatly reduced in hepatocytes. Moreover, HCC cells have evolved to transfer oncogenic ncRNAs to recipient cells via exosomes, steadily transmitting biological signals and promoting tumor growth [32, 33]. Similarly, it has been reported that miRNAs may also regulate the activity and function of p53 by directly targeting UBE4B [34]. However, there is no clear evidence for a relationship between hsa-miR-22-3p and tumorigenesis. Our results demonstrated a tumor process between UBE4B and HCC. Therefore, we propose a novel regulatory axis for targeted drugs, namely UBE4B-hsa-miR-22-3p-FGD5-AS1/LINC00858/SNHG16. Moreover, we confirmed that the mRNA expression of FGD5-AS1, LINC00858, SNHG16, and UBE4B was higher in HCC cell lines than that in normal hepatocyte lines, and the expression of hsa-miR-22-3p mRNA showed a decreasing trend.
Non-coding RNAs are important components of exosomes, and can regulate the TME and promote HCC progression, including development, progression, infiltration, metastasis, and angiogenesis [35]. These are traits of tumor immuno-infiltration and immune escape. Moreover, oncogenic ncRNAs from exosomes may also suppress the activity of immunosuppressive cells such as T-effector cells, ultimately leading to local or systemic immunosuppression. Our study showed the same mechanism, as evidenced by all the immune cells analyzed in HCC, including B cells, CD8+ T cells, CD4+ T cells, macrophages, neutrophils, and dendritic cells. Furthermore, UBE4B expression was significantly and positively correlated with most immune cell biomarkers in HCC.
The degree of the immune response is regulated by the balance between the immunoinhibitor and immunostimulator pathways. These receptors and ligands are known as immune checkpoints [36]. Our study showed that UBE4B is significantly associated with immunoinhibitors and immunostimulators in HCC. The expression of UBE4B was significantly associated with many chemokines and chemokine receptors, suggesting that UBE4B may be associated with immune regulation in HCC and that these molecules could be potential immunotherapeutic targets for UBE4B in HCC.
The current study had some limitations. First, most data are based on online databases that are constantly updated and expanded, which may affect the results of the study. Second, this study requires additional in vivo and in vitro experiments to examine the functional mechanisms of the ceRNA network. Third, these results need to be validated in large-scale clinical trials.
Conclusions
Taken together, our results suggest that UBE4B expression was increased in HCC and was correlated with a poor prognosis in patients with HCC. UBE4B expression was positively correlated with immune cell infiltration and immunomodulators, chemokines, and their receptors, further suggesting that UBE4B plays a carcinogenic role by modulating immune processes. Additionally, we identified the indispensable role of the UBE4B-hsa-miR-22-3p-FGD5-AS1/LINC00858/SNHG16 regulatory axis in HCC. These findings will aid in understanding the relevant functions of UBE4B and provide new strategies for drug development and exploration of prognosis-related biomarkers.
Author Contributions
X. Shao, J. Zhu, and Y. Shi designed this study. H. Fang and J. Chen collected samples. Y. Zhang, J. Wang, and H. Jian performed the experiments and analyzed the results. S. Lan and F. Jiang drafted the manuscript. F. Zhong, Y. Zhang, and C. Cao provided the conceptualization and methodology. All authors contributed to the manuscript and approved the submitted version.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Funding
This study was supported by Scientific Research Fund of Anhui Medical University (grant number 2020xkj058), 2021 Anhui Provincial Health Commission Scientific Research Project (grant number AHWJ2021b097), 2022 Provincial Key Research and Development Plan Projects (grant number 2022e07020050), Jiangsu Province Foreword Leading Technology Fundamental Research Project (grant number BK20212012), National Natural Science Foundation of China (grant number 62141109), Jiangsu Province Social Development Project (grant number BE2022812) and 2019 Jiaxing Medical Key Discipline Construction Project (grant number 2019-SZ-009).
References
- 1. McGlynn KA, Petrick JL, El-Serag HB. Epidemiology of Hepatocellular Carcinoma. Hepatology. 2021 (Suppl 1); 73:4–13. https://doi.org/10.1002/hep.31288 [PubMed]
- 2. Llovet JM, Kelley RK, Villanueva A, Singal AG, Pikarsky E, Roayaie S, Lencioni R, Koike K, Zucman-Rossi J, Finn RS. Hepatocellular carcinoma. Nat Rev Dis Primers. 2021; 7:6. https://doi.org/10.1038/s41572-020-00240-3 [PubMed]
- 3. Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, Bray F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin. 2021; 71:209–49. https://doi.org/10.3322/caac.21660 [PubMed]
- 4. Kulik L, El-Serag HB. Epidemiology and Management of Hepatocellular Carcinoma. Gastroenterology. 2019; 156:477–91.e1. https://doi.org/10.1053/j.gastro.2018.08.065 [PubMed]
- 5. Anwanwan D, Singh SK, Singh S, Saikam V, Singh R. Challenges in liver cancer and possible treatment approaches. Biochim Biophys Acta Rev Cancer. 2020; 1873:188314. https://doi.org/10.1016/j.bbcan.2019.188314 [PubMed]
- 6. Malik A, Thanekar U, Amarachintha S, Mourya R, Nalluri S, Bondoc A, Shivakumar P. “Complimenting the Complement”: Mechanistic Insights and Opportunities for Therapeutics in Hepatocellular Carcinoma. Front Oncol. 2021; 10:627701. https://doi.org/10.3389/fonc.2020.627701 [PubMed]
- 7. European Association for the Study of the Liver. Electronic address:
[email protected] , and European Association for the Study of the Liver. EASL Clinical Practice Guidelines: Management of hepatocellular carcinoma. J Hepatol. 2018; 69:182–236. https://doi.org/10.1016/j.jhep.2018.03.019 [PubMed] - 8. Huang J, Dong J, Shi X, Chen Z, Cui Y, Liu X, Ye M, Li L. Dual-Functional Titanium(IV) Immobilized Metal Affinity Chromatography Approach for Enabling Large-Scale Profiling of Protein Mannose-6-Phosphate Glycosylation and Revealing Its Predominant Substrates. Anal Chem. 2019; 91:11589–97. https://doi.org/10.1021/acs.analchem.9b01698 [PubMed]
- 9. Han ZJ, Feng YH, Gu BH, Li YM, Chen H. The post-translational modification, SUMOylation, and cancer (Review). Int J Oncol. 2018; 52:1081–94. https://doi.org/10.3892/ijo.2018.4280 [PubMed]
- 10. Hickey CM, Xie Y, Hochstrasser M. DNA binding by the MATα2 transcription factor controls its access to alternative ubiquitin-modification pathways. Mol Biol Cell. 2018; 29:542–56. https://doi.org/10.1091/mbc.E17-10-0589 [PubMed]
- 11. Qu Y, Dang S, Hou P. Gene methylation in gastric cancer. Clin Chim Acta. 2013; 424:53–65. https://doi.org/10.1016/j.cca.2013.05.002 [PubMed]
- 12. Hatakeyama S, Nakayama KI. U-box proteins as a new family of ubiquitin ligases. Biochem Biophys Res Commun. 2003; 302:635–45. https://doi.org/10.1016/s0006-291x(03)00245-6 [PubMed]
- 13. Memarzadeh K, Savage DJ, Bean AJ. Low UBE4B expression increases sensitivity of chemoresistant neuroblastoma cells to EGFR and STAT5 inhibition. Cancer Biol Ther. 2019; 20:1416–29. https://doi.org/10.1080/15384047.2019.1647049 [PubMed]
- 14. Wu H, Pomeroy SL, Ferreira M, Teider N, Mariani J, Nakayama KI, Hatakeyama S, Tron VA, Saltibus LF, Spyracopoulos L, Leng RP. UBE4B promotes Hdm2-mediated degradation of the tumor suppressor p53. Nat Med. 2011; 17:347–55. https://doi.org/10.1038/nm.2283 [PubMed]
- 15. Weng C, Chen Y, Wu Y, Liu X, Mao H, Fang X, Li B, Wang L, Guan M, Liu G, Lu L, Yuan Y. Silencing UBE4B induces nasopharyngeal carcinoma apoptosis through the activation of caspase3 and p53. Onco Targets Ther. 2019; 12:2553–61. https://doi.org/10.2147/OTT.S196132 [PubMed]
- 16. Huang XQ, Hao S, Zhou ZQ, Huang B, Fang JY, Tang Y, Zhang JH, Xia JC. The Roles of Ubiquitination Factor E4B (UBE4B) in the Postoperative Prognosis of Patients with Renal Cell Carcinoma and in Renal Tumor Cells Growth and Metastasis. Onco Targets Ther. 2020; 13:185–97. https://doi.org/10.2147/OTT.S229577 [PubMed]
- 17. Zhang Y, Lv Y, Zhang Y, Gao H. Regulation of p53 level by UBE4B in breast cancer. PLoS One. 2014; 9:e90154. https://doi.org/10.1371/journal.pone.0090154 [PubMed]
- 18. Li T, Fan J, Wang B, Traugh N, Chen Q, Liu JS, Li B, Liu XS. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017; 77:e108–10. https://doi.org/10.1158/0008-5472.CAN-17-0307 [PubMed]
- 19. Tang Z, Li C, Kang B, Gao G, Li C, Zhang Z. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017; 45:W98–102. https://doi.org/10.1093/nar/gkx247 [PubMed]
- 20. Ru B, Wong CN, Tong Y, Zhong JY, Zhong SS, Wu WC, Chu KC, Wong CY, Lau CY, Chen I, Chan NW, Zhang J. TISIDB: an integrated repository portal for tumor-immune system interactions. Bioinformatics. 2019; 35:4200–2. https://doi.org/10.1093/bioinformatics/btz210 [PubMed]
- 21. Qi X, Zhang DH, Wu N, Xiao JH, Wang X, Ma W. ceRNA in cancer: possible functions and clinical implications. J Med Genet. 2015; 52:710–18. https://doi.org/10.1136/jmedgenet-2015-103334 [PubMed]
- 22. Li W, Wang B, Dong S, Xu C, Song Y, Qiao X, Xu X, Huang M, Yin C. A Novel Nomogram for Prediction and Evaluation of Lymphatic Metastasis in Patients With Renal Cell Carcinoma. Front Oncol. 2022; 12:851552. https://doi.org/10.3389/fonc.2022.851552 [PubMed]
- 23. Vautrot V, Bentayeb H, Causse S, Garrido C, Gobbo J. Tumor-Derived Exosomes: Hidden Players in PD-1/PD-L1 Resistance. Cancers (Basel). 2021; 13:4537. https://doi.org/10.3390/cancers13184537 [PubMed]
- 24. Hughes CE, Nibbs RJ. A guide to chemokines and their receptors. FEBS J. 2018; 285:2944–71. https://doi.org/10.1111/febs.14466 [PubMed]
- 25. Woodfield SE, Guo RJ, Liu Y, Major AM, Hollingsworth EF, Indiviglio S, Whittle SB, Mo Q, Bean AJ, Ittmann M, Lopez-Terrada D, Zage PE. Neuroblastoma patient outcomes, tumor differentiation, and ERK activation are correlated with expression levels of the ubiquitin ligase UBE4B. Genes Cancer. 2016; 7:13–26. https://doi.org/10.18632/genesandcancer.97 [PubMed]
- 26. Zhang XF, Pan QZ, Pan K, Weng DS, Wang QJ, Zhao JJ, He J, Liu Q, Wang DD, Jiang SS, Zheng HX, Lv L, Chen CL, et al. Expression and prognostic role of ubiquitination factor E4B in primary hepatocellular carcinoma. Mol Carcinog. 2016; 55:64–76. https://doi.org/10.1002/mc.22259 [PubMed]
- 27. Koleckova M, Ehrmann J, Bouchal J, Janikova M, Brisudova A, Srovnal J, Staffova K, Svoboda M, Slaby O, Radova L, Vomackova K, Melichar B, Veverkova L, Kolar Z. Epithelial to mesenchymal transition and microRNA expression are associated with spindle and apocrine cell morphology in triple-negative breast cancer. Sci Rep. 2021; 11:5145. https://doi.org/10.1038/s41598-021-84350-2 [PubMed]
- 28. Mukherjee S, Akbar I, Bhagat R, Hazra B, Bhattacharyya A, Seth P, Roy D, Basu A. Identification and Classification of Hubs in microRNA Target Gene Networks in Human Neural Stem/Progenitor Cells following Japanese Encephalitis Virus Infection. mSphere. 2019; 4:e00588–19. https://doi.org/10.1128/mSphere.00588-19 [PubMed]
- 29. Koźniewski K, Wąsowski M, Jonas MI, Lisik W, Jonas M, Binda A, Jaworski P, Tarnowski W, Noszczyk B, Puzianowska-Kuźnicka M, Kuryłowicz A. Epigenetic Regulation of Estrogen Receptor Genes’ Expressions in Adipose Tissue in the Course of Obesity. Int J Mol Sci. 2022; 23:5989. https://doi.org/10.3390/ijms23115989 [PubMed]
- 30. Bhatt P, d’Avout C, Kane NS, Borowiec JA, Saxena A. Specific domains of nucleolin interact with Hdm2 and antagonize Hdm2-mediated p53 ubiquitination. FEBS J. 2012; 279:370–83. https://doi.org/10.1111/j.1742-4658.2011.08430.x [PubMed]
- 31. Du C, Wu H, Leng RP. UBE4B targets phosphorylated p53 at serines 15 and 392 for degradation. Oncotarget. 2016; 7:2823–36. https://doi.org/10.18632/oncotarget.6555 [PubMed]
- 32. Mashouri L, Yousefi H, Aref AR, Ahadi AM, Molaei F, Alahari SK. Exosomes: composition, biogenesis, and mechanisms in cancer metastasis and drug resistance. Mol Cancer. 2019; 18:75. https://doi.org/10.1186/s12943-019-0991-5 [PubMed]
- 33. Sasaki R, Kanda T, Yokosuka O, Kato N, Matsuoka S, Moriyama M. Exosomes and Hepatocellular Carcinoma: From Bench to Bedside. Int J Mol Sci. 2019; 20:1406. https://doi.org/10.3390/ijms20061406 [PubMed]
- 34. Wang B, Wu H, Chai C, Lewis J, Pichiorri F, Eisenstat DD, Pomeroy SL, Leng RP. MicroRNA-1301 suppresses tumor cell migration and invasion by targeting the p53/UBE4B pathway in multiple human cancer cells. Cancer Lett. 2017; 401:20–32. https://doi.org/10.1016/j.canlet.2017.04.038 [PubMed]
- 35. Liu J, Fan L, Yu H, Zhang J, He Y, Feng D, Wang F, Li X, Liu Q, Li Y, Guo Z, Gao B, Wei W, et al. Endoplasmic Reticulum Stress Causes Liver Cancer Cells to Release Exosomal miR-23a-3p and Up-regulate Programmed Death Ligand 1 Expression in Macrophages. Hepatology. 2019; 70:241–58. https://doi.org/10.1002/hep.30607 [PubMed]
- 36. Topalian SL, Taube JM, Anders RA, Pardoll DM. Mechanism-driven biomarkers to guide immune checkpoint blockade in cancer therapy. Nat Rev Cancer. 2016; 16:275–87. https://doi.org/10.1038/nrc.2016.36 [PubMed]