



Introduction
The aging population
Aging is the gradual process of organismal deterioration which is associated with a multitude of age-related disorders and diseases that make one wonder if aging itself is a disease that needs to be addressed [1]. A shadow is cast on the benefits of longevity if the elderly are faced with the possibility of a decline in their quality of life. The world currently has over 700 million people who are over the age of 65, a number that is projected to grow rapidly in the near future [2]. As advancing age is strongly correlated to decreased quality of life and increased risk of several age-related diseases [3], these demographics seem more dismal in prospering countries, with the USA and the UK having about 16–18% of their population over the age of 65 [4, 5]. With the life expectancy of most Western countries steadily increasing, majority of people are expected to spend at least 2 decades, or 25% of their life, over the age of 65, when they are prone to acquiring various age-related morbidities [6, 7]. The silver lining to this otherwise tragic situation is that results from recent studies indicate that the aging process and the pace of organismal deterioration is malleable and can be influenced greatly by physiological, genetic, dietary and pharmaceutical interventions [8–16].
The aging immune system
The immune system is a complex network of cells and tissues working in coalition to maintain the health of an organism. It not only clears foreign pathogens, but also helps to maintain the integrity of the organism by clearing away dead or dysfunctional cells [17–22]. Due to the immune system’s complexity and intricacy, 7% of the genes from the human genome are allocated exclusively for its functioning and maintenance [23].
Like any other system, the immune system changes with age and experiences gradual deterioration. Improving our understanding of this phenomenon is of great significance because the medical and scientific advancements that have facilitated the unprecedented increase in average human lifespan have been unable to significantly increase the human healthspan [24]. Because of this, we have a rapidly increasing aging population in a world where there is a substantial risk of steep decline in quality of life with age.
Age-associated deterioration and dysfunction of the immune system leads to the establishment of an incompetent immune response against invading pathogens [25, 26]. This could partially provide an explanation for the age-dependent increase of mortality in patients suffering from infections like influenza [27], with people older than 65 accounting for more than 90% of the influenza-associated annual deaths [28]. Furthermore, the aged immune system elicits an inadequate response to vaccines, leaving the elderly susceptible to pathogens despite being vaccinated against them [29, 30]. This is especially poignant in the wake of an ongoing pandemic where the mortality rate is disproportionately high in the elderly [31].
Aging of the immune system is also one of the major factors that accelerates the deterioration of an organism, as its dysfunction not only fails to elicit a strong immune response against invading pathogens but also drives the accumulation of undesirable and malfunctioning cells [25, 32–36]. In some cases the aging immune system also develops an affinity for attacking self-antigens, leading to autoimmunity-associated disorders [37, 38].
In recent years, there have been many studies that have broadened our understanding of the aging immune system and immunosenescence (the gradual deterioration of the immune system with age) from the perspective of genetics, nutrition, physiology, and molecular biology [39–42]. Despite this assimilation of knowledge, a complete understanding of the dynamics of this process is lacking.
Within a systemic context, the age-related changes and adversities in any organ system arise from a complex crosstalk between different cells and processes of the body. By virtue of the way that research studies are designed and funded, many aspects of this complexity are often overlooked. In this review, we will discuss one such interaction, between cellular senescence and the immune system with a focus on the accumulation of SnCs in the lymphoid organs of the aging body, which is greatly understudied and underappreciated.
Cellular senescence
Initially described in 1961, cellular senescence is the phenomenon by which cells cease to divide despite the availability of adequate growth factors [43]. It was later established that upon encountering certain types of stress and irreparable damage, cells tend to enter a stable cell cycle arrest [44]. From an evolutionary perspective, this is widely considered to be a protective mechanism to prevent the stressed and damaged cells from becoming deleterious to the body.
Like most things optimized by evolution, cellular senescence is not of much concern to the younger body capable of reproduction while the older body, past its reproductive prime, is adversely affected by it. The fitness benefits that cellular senescence provides to younger, reproductively active animals, such as preventing cancer [45], mitigating the progression of fibrosis [46–48] and promoting optimal wound healing [49], have helped the phenomenon survive the arduous tests of natural selection over the millennia. Unfortunately, in almost an antagonistically pleiotropic manner, accumulation of SnCs is very detrimental to the older body [50]. Specifically, SnCs secrete various factors classified together as senescence-associated secretory phenotype (SASP) which cause instability and dysfunction in their surrounding environment [51]. Both SnCs and SASP factors have been implicated in many of the age-related deteriorations, dysfunctions and diseases including but not limited to frailty, hypertrophy of tissue, stem-cell exhaustion, bystander effect mediated senescent cell accumulation, and cancer [51–63].
The interactions between SnCs and the immune system run in both directions, with the immune system surveilling and clearing the SnCs; while the SnCs frequently impede the function, and in some contexts, generation of immune cells. In young and healthy individuals, the immune system can rapidly clear SnCs after their induction, which prevents them from significantly accumulating and causing adverse effects [18, 64]. In older individuals, this turnover is slow and leads to the accumulation of SnCs [34]. Ovadya et al. demonstrated that accumulation of SnCs is accelerated upon impaired immune surveillance [32]. Since advancing age is associated with impairment in immune function [65], the decline in the turnover of SnCs with age can, at least partially, be attributed to this impediment. Despite multiple studies demonstrating various mechanisms via which SnCs could evade immune clearance [66, 67], the impact of aging on immune evasion of SnCs is not yet completely understood. Of note, SnCs have been shown to cause stem cell exhaustion and dysfunction [62, 68–72]. This is of great relevance and importance to the topic of immunosenescence because senescence, exhaustion and dysfunction of hematopoietic stem cells (HSCs), causes myeloid skewing and a decrease in the production of immune cells which may be one of the underlying causes of age-related immunosenescence.
With many more possible domains of interaction between cellular senescence and the immune system, as seen in (Figure 1), this review will discuss literature that states or suggests the presence of this interaction, with a focus on cellular senescence in the lymphoid organs, and raises questions that need to be answered to strengthen the foundation of the role of cellular senescence in immunosenescence.
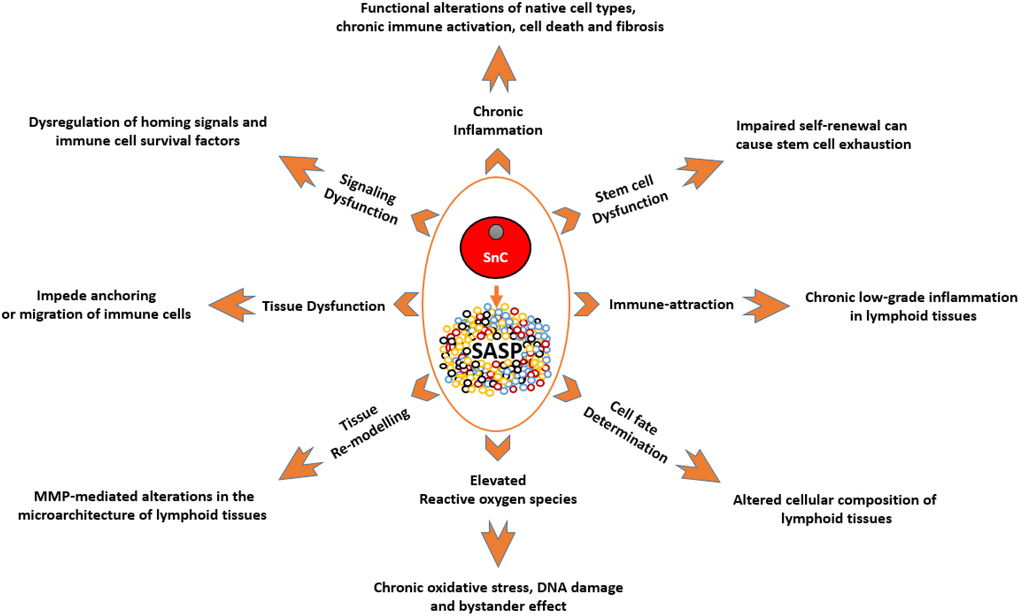
Figure 1. A depiction of the known effects of SnCs and SASP on different cell types and tissues, and how they are relevant to the immune system. SnCs possess altered morphology and surface markers and usually fail to perform the tasks of their non-senescence counterparts. This makes them the dysfunctional units of a tissue which can impede normal functions such as, immune cell priming and transmigration. MMPs produced by SnCs can modify the surrounding matrix and alter the microarchitecture of the lymphoid organs. As these organs are precisely organized into zones with specialized functions, such micro-architectural alterations can lead to dysfunction. SASP produced by SnCs can act as a chemoattractant to immune cells which can lead to unresolved chronic inflammation in tissues. SASP by itself can be inflammatory which can adversely impact neighboring cells. This chronic unresolved inflammation can lead to pathological conditions like fibrosis and neoplasia. SASP-mediated signaling and ROS-mediated oxidative stress can impair clonogenicity and functionality of HSCs, immune cells and other supporting cells of the immune system. SnCs and SASP can alter the expression profile of supporting cells leading to the dysregulation of homing signals required for proper localization of immune cells, and survival factors required for the endurance of certain immune cells. SnCs, by means of SASP, can influence the cell fate of differentiating cells and in some cases, cause the accumulation of adipocytes in the lymphoid organs. Abbreviations: SnC: Senescent cell; SASP: Senescence associated secretory phenotype; MMPs: Matrix metalloproteases; ROS: Reactive Oxygen Species; HSC: Hematopoietic stem cell.
Cellular Senescence in the Organs of the Immune System
Bone marrow
Bone marrow is a spongy tissue residing in the core of vertebrae, skull and long bones. It is the home of HSCs which give rise to most of the immune cells [73]. HSCs are self-renewing pluripotent cells that can generate the entire hematopoietic system.
With increasing age, the bone marrow microenvironment changes dramatically. With advancing age, HSC number increases, while their functionality, including self-renewal and clonogenicity declines. These changes are accompanied by myeloid skewing, elevated adipogenesis in the bone marrow, and alterations in the bone marrow niche [74–78]. Along with the prevalence of significantly more apoptotic cells, bone marrow cellularity (volume occupied by HSCs) decreases significantly with age reaching values lower than 40% [79]. A graphic depiction of the aged bone marrow microenvironment is illustrated in (Figure 2).
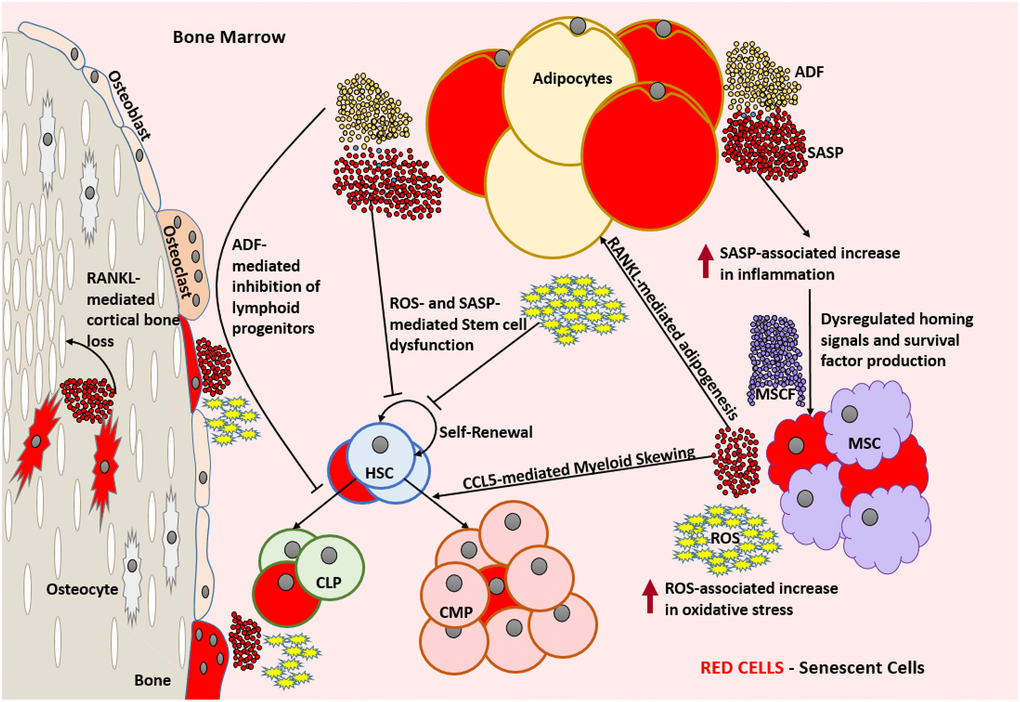
Figure 2. Aged bone marrow microenvironment with accumulated SnCs is not conducive for its normal functionality. SASP and ROS mediate dysfunction and DNA damage in HSCs, respectively and lead to a change in the HSC repertoire and exhaustion of the functional HSC reservoir. RANKL mediates the accumulation of adipocytes that produce ADFs. CCL5 and ADFs mediate the establishment of myeloid skewing in HSCs. SASP mediated inflammation can dysregulate the adequate production of homing signals and survival factors by the MSCs which can lead to the depletion of selective immune cell types. The increased ROS and SASP mediated inflammation causes damage to the surrounding cells and induces senescence by means of the bystander effect. SnCs such as osteocytes can produce SASP that is detrimental to the bone housing which encloses them. In the absence of rapid clearance of SnCs, this becomes a self-perpetuating cycle of dysfunction and damage causing severe immunosenescence. Abbreviations: SnC: Senescent cell; SASP: Senescence associated secretory phenotype; ROS: Reactive Oxygen Species; HSC: Hematopoietic stem cell; CLP: Common lymphoid progenitor; CMP: Common myeloid progenitor; MSC: Mesenchymal stem cell; MCSF: Mesenchymal stem cell derived factors; ADF: Adipocyte derived factors; CCL5: Chemokine Ligand 5; RANKL: Receptor activator of nuclear factor kappa-Β ligand.
Myeloid skewing of HSCs with aging may be in part attributable to the aged bone marrow microenvironment, as even young HSCs develop a myeloid bias upon being transplanted into old mice [80, 81]. It has been suggested that chemokine ligand 5 (CCL5) is a major factor that drives myeloid skewing of HSCs with advancing age. Over expression of CCL5 causes a decrease in pro-lymphoid transcription factors and T-cell differentiation, while genetically knocking out CCL5 prevents myeloid skewing in mice [82]. Age-related accumulation of adipocytes in the bone marrow has been attributed to the increased expression of receptor activator of nuclear factor kappa-B ligand (RANKL) [83]. These bone marrow adipocytes in-turn produce an array of factors that have been shown to affect hematopoiesis and skew it towards myeloid lineage [84–88].
The accumulation of various p16INK4a positive cells [89–91], SASP factor (like CCL5 and RANKL) generating cells [91–94] in aged bone marrow, along with increased number of cells harboring DNA damage and elevated ROS [95, 96], suggests that age-dependent bone marrow changes can be in part attributed to the accumulation of SnCs.
Based on data showing that the expression profile of adipocytes resembles the SASP profile of SnCs [97], it is likely that a great proportion of these adipocytes are senescent. This became evident after a study where clearance of SnCs in INK-ATTAC mice, a genetically altered model that clears cells expressing p16INK4a, showed a significant reduction in the number, size, and tissue volume of bone marrow adipocytes [98]. Other studies have also shown that, despite the structural and functional support provided by adipocytes, they adversely influence the hematopoietic environment [99, 100]. However, whether this is completely attributable to senescent adipocytes and their SASP is yet to be determined.
A recent study implicated the senescence of bone marrow-derived mesenchymal stem cells (BM-MSCs) in the age-associated dysfunction of HSCs, in humans. This study revealed that a significantly higher portion of senescent MSCs were seen in the bone marrow explants of the elderly when compared to their younger cohorts. This was established by showing increased accumulation of cells with DNA damage, elevated ROS and SASP expression. They also showed that the functionality and clonogenicity of young HSCs were impaired when exposed to factors generated by these MSCs [95]. The inflammatory environment, created by SASP of these SnCs, can alter the expression profile of normal MSCs to dysregulate the expression of factors necessary for lymphocyte survival [101–105].
Along with the cell-extrinsic causes for stem cell aging, older HSCs show an accumulation of senescence in association with increased DNA damage and telomere attrition, along with having an increased risk of undergoing an inflammatory cell death known as pyroptosis [68, 106]. Reactive oxygen species (ROS) produced by SnCs play a key role in the bystander effect [107]. ROS produced by SnCs in the bone marrow environment can cause DNA breaks in HSCs. This agrees with the finding that aged HSCs harbor more DNA damage compared to their younger counterparts [108]. As the DNA damage repair mechanism is not robust and quite error prone in the quiescent HSCs [109], the constant oxidative stress-induced DNA damage can progressively deplete and alter the functional HSC repertoire with increasing age [110].
Direct evidence for the adverse role of cellular senescence in modulating HSC function during aging was provided by demonstrating that knocking out p16INK4a conserved HSC functionality and stress tolerance with age [68]. A more recent study from our lab has shown that clearing SnCs rejuvenated the aging HSC repertoire by reducing myeloid skewing and improving clonogenicity significantly in mice [63].
Thymus
The thymus is a primary lymphoid organ located behind the breastbone and above the heart, within which T-cells mature. In an evolutionarily conserved manner, most vertebrates experience an age-associated thymic involution, which is characterized by atrophy and the development of cavities. An age-dependent alteration in thymic cellularity can be seen, with most functional cells getting replaced by fibroblasts, fat cells and senescent cells [111–116].
Thymic atrophy is associated with the reduced turnover of new T-cells [117], a constricted T-cell receptor repertoire [118] and the production of higher autoreactive T-cells that could lead to autoimmunity [119]. As depicted in (Figure 3) these are characteristic features of immunosenescence that play an important role in age-associated impaired T-cell function [120].
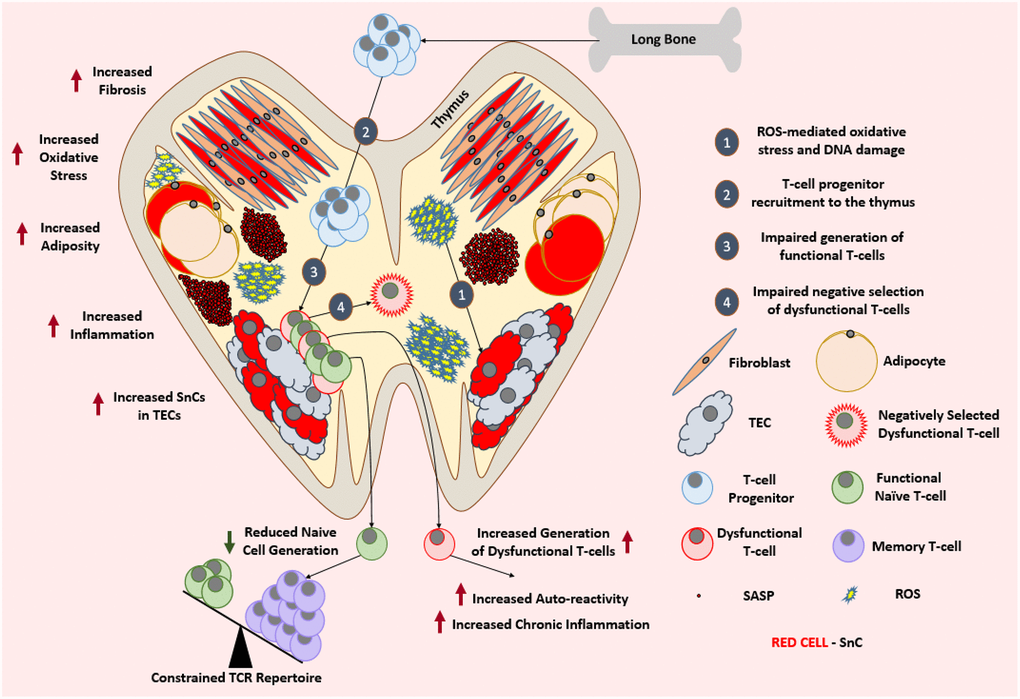
Figure 3. Aged thymus is dysfunctional. With advancing age, thymus loses its cellularity while accumulating adipocytes and fibroblasts. Aged thymus develops an inflammatory environment with high levels of oxidative stress. This is evident by the accumulation of senescent TECs with elevated markers of DNA damage and oxidative stress. Despite the adequate recruitment of T-cell progenitors, aged thymus generates inadequate number of naïve T-cells which leads to the age-associated depletion of TCR repertoire and ultimately a change in the immune cell landscape. Due to the impaired negative selection of dysfunctional T-cells, the aged thymus shows an increase in the output of dysfunctional and autoreactive T-cells leading to the establishment of low-grade chronic inflammation. Abbreviations: SnC: Senescent cell; SASP: Senescence associated secretory phenotype; ROS: Reactive Oxygen Species; TEC: Thymic epithelial cell; TCR: T-cell receptor.
Thymic epithelial cells (TECs) from adult human thymus stained positive for senescence- associated beta galactosidase (SA-βGal) and the thymic tissues from these adults also strongly stained positive for markers of oxidative DNA damage such as γH2AX and 8-oxoguanine [121]. A similar finding of high γH2AX staining was seen in the thymus of old mice, which was indicative of DNA damage and cellular senescence [111]. This also correlated with the increased inflammatory environment of the aged thymus seen in humans [122]. Despite the abundance of evidence suggesting accumulation of SnCs in atrophied thymus, whether cellular senescence plays a causal role in thymic involution needs to be further studied, as the accumulation of SnCs could be a consequence of thymic involution. But the possibility of a causal involvement of SnCs and their SASP seems likely because the administration of IL-6, a known SASP factor, has been shown to induce thymic atrophy [122]. In addition, increased oxidative stress and DNA damage in the stromal cells, especially TECs, has also been shown to accelerate thymic aging [123].
With the existing knowledge that TECs play a crucial role in the positive and negative selection of maturing T-cells [124], the role of senescent TECs in the thymic environment should also be explored in the context of positive and negative selection of T-cells. For example, it has yet to be determined whether the interaction of the developing T-cells with SnCs of the thymus play a role in the development of T-cells with auto-reactivity.
Interestingly, the recruitment of T-cell progenitors to the thymus is similar between young and old mice [125]. The reduced T-cell output has been attributed to the defective microenvironment of the thymus and other secondary lymphoid organs [125–127]. Though there is a significant functional decline in thymic activity, the aged thymus still retains a portion of its function [128], which leads us to believe that the therapeutic clearance of SnCs could help to restore thymic function in the elderly. Thymic regeneration strategies so far have largely failed to improve the production of functional of T-cells, in part due to the lack of a systemic approach, because rejuvenating the thymus alone still leaves the secondary lymphoid organs too impaired to support the naïve T-cells being produced [127, 129].
It would be intriguing to replicate these studies with a senolytic combinatorial therapy to see how it changes the outcome. It should be a promising venture, because caloric restriction, a dietary intervention known to reduce cellular senescence [130] and SASP [131, 132], has been shown to delay thymic involution and mitigate thymic adipogenesis [133].
Spleen
The spleen is a secondary lymphoid organ that acts as a blood filter to remove damaged red blood cells. It plays a crucial role in maintaining the optimal populations of white blood cells and platelets. The spleen can detect pathogenic invaders in the blood and mobilize the immune system to fight against the pathogens [134].
With advancing age, the cellularity and microarchitecture of the spleen changes significantly accompanied by altered localization of various cells [135]. The distinct demarcation of T-cell and B-cell regions within the white pulp becomes obscure with advancing age. Also, an alteration in the organization and function of stromal cells, marginal zone macrophages and marginal metallophilic macrophages can be seen [136]. An accumulation of SnCs with advancing age has been demonstrated to happen in the spleen. This was shown not only by means of elevated expression of p16INK4a and SASP factors, but also by means of cell accumulation with elevated DNA damage [50, 137]. It has also been shown that the stromal cell populations of the aged spleen, exhibit an upregulated expression of IL-6, a SASP factor, implying that at least a proportion of these cells could be senescent [138].
Age-dependent changes in the splenic microenvironment impair the phagocytic capacity of macrophages in the marginal zone. While the phagocytic capacity of macrophages from the aged spleen seemed to be less efficient in vivo, their in vitro phagocytic capacity was similar to those from young mice [139]. Interestingly, induction of SnCs accumulation in the spleen after radiation has been shown to impart similar functional impairments to splenic macrophages in mice, and the clearance of such SnCs was able to restore macrophage function [140]. Microenvironment-dependent dysfunction and impaired migration of B-cells can also be seen in the aged spleen [141]. Even B-cells originating from young HSCs in an aged recipient showed signs of dysfunction, providing support to the idea that B-cell dysfunction is mainly attributable to the aged splenic environment [135].
Splenic priming of T-cells is a crucial step in the establishment of an appropriate T-cell response [142]. It is known that the senescent splenic environment impairs the recruitment of T-cells to the spleen. In addition, as depicted in (Figure 4), the microenvironment-mediated impairment of the functionality of antigen-presenting cells such as B-cells, macrophages and dendritic cells (DCs) in the aged spleen may explain why even T-cells originating from young HSCs were dysfunctional and showed a delayed response to stimulation in an aged splenic microenvironment [143, 144].
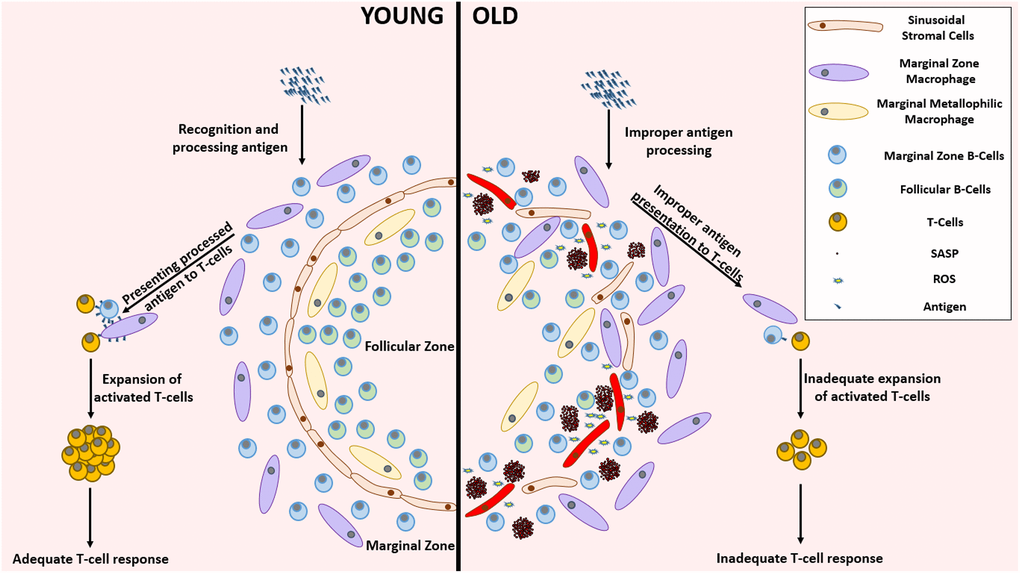
Figure 4. Remarkable differences between the young and aged splenic environment. With advancing age, the stromal cells in the lining of sinuses, that demarcate follicular zone from the marginal zone, become less organized accompanied with an altered localization of various cell types. The inflammatory environment created by the accumulation of SnCs impairs the functionality of several cells residing in the spleen. This functional impairment mediated improper antigen presenting capabilities lead to the establishment of an inadequate T-cell response against pathogenic invasion. Abbreviations: SnC: Senescent cell; SASP: Senescence associated secretory phenotype; ROS: Reactive Oxygen Species.
Though not deeply explored in these studies, it is apparent that the splenic environment of the old mice is not conducive for the proper functionality of various immune cells. Apart from SASP-mediated micro-environmental alterations, SnCs, by virtue of their altered morphology, can imbue structural alterations to the aged spleen. With senolytics [14, 15, 145–148] and senostatics [149] becoming more accessible, further insights into SASP-independent mechanisms of SnCs involvement in immunosenescence should be explored.
Lymph nodes
Lymph nodes are small bulbous structures that form a crucial part of the lymphatic system along with the lymphatic vessels. They filter the lymph fluid obtained from the surrounding tissues before it re-enters the blood stream [150].
Lymph nodes house various immune cells including T-cells, B-cells and DCs and play an essential role in establishing a strong immune response [151–154]. With advancing age, there is a significant decline in the number, integrity, and functionality of lymph nodes [135, 155–158]. Alterations in cellularity and functionality of different cell types of lymph nodes have been shown to occur with advancing age (reviewed here [158]). Increased adiposity and fibrosis have also been described in lymph nodes of patients older than 60 years [155, 156].
It has been speculated that lymphatic endothelial cells and high endothelial venules of the lymph nodes show signs of aging similar to that of the vascular system. This includes altered permeability, accumulation of SnCs, and increased inflammation, which could act as causal factors that adversely affect the migration and recruitment of immune cells like naïve T-cells [158]. It has also been shown that the age-dependent increase in the level of prostaglandin-2 in the lungs inhibits the migration of DCs to the draining lymph nodes, leading to the establishment of an improper T-cell response to viral infections like SARS-CoV [159]. This is interesting since prostaglandin production is upregulated in SnCs [160], and provides evidence on how cellular senescence in other organs can indirectly impact the function of lymph nodes.
Stromal cells from aged lymph nodes have reduced replicative potential upon stimulation [161, 162] and were unable to support naïve T-cell homeostasis [127]. Though not explored as a possibility in these studies, this could be an indication that at least a portion of these stromal are senescent. Another interesting study sheds light on the role of chemokine ligand 2 (CCL2) produced by the stromal cells of lymph nodes in the mitigation of antibody response [163]. Despite this being an important function that prevents the establishment of unnecessary germinal centers in the absence of an antigen, CCL2 is a SASP factor, which raises the question of whether senescent stromal cells that perpetually produce CCL2 are responsible for the age-dependent impairment of lymph nodes to support germinal centers [157].
It seems highly likely that cellular senescence is involved in this age-related lymph node deterioration. Further studies exploring the presence of SnCs in the aged lymph nodes and their role in lymph node-mediated immune response are needed.
Mucosa associated lymphoid tissue
Mucosa-associated lymphoid tissue (MALT) is a part of the immune system that localizes on the surface of the mucosal tissues. Depending on their location, MALT is classified into different types, such as inducible bronchus-associated lymphoid tissue (iBALT) [164, 165], conjunctiva-associated lymphoid tissue (CALT) [166, 167], larynx-associated lymphoid tissue (LALT) [168] and inducible skin-associated lymphoid tissue (SALT) [169, 170]. The most commonly studied MALT representatives are nasopharynx-associated lymphoid tissue (NALT) [171, 172] and gut-associated lymphoid tissue (GALT) [173].
In humans, the adenoids of the nasopharynx, tonsils of oropharynx, and a few more lymph nodes in the region form the Waldeyer’s ring [174, 175]. They are considered to be a part of the MALT and are analogous to the NALT in rodents [172]. They are crucial for immunization through intranasal vaccination [176]. Similarly, GALT is comprised of Peyer’s patches, mesenteric lymph nodes (MLNs) and isolated lymphoid follicles (ILFs) [177].
The MALT functions in a complex manner (reviewed here [178]), which is known to be affected by the process of aging, as seen in mice by the age-dependent reduction in the establishment of oral tolerance to novel antigens [179]. This deterioration varies regionally, with NALT conserving its functionality for longer than GALT, making nasal immunizations an attractive alternative for vaccinating the elderly [180, 181].
Though cellular senescence has been shown to be present in the tonsils of patients with tonsillitis and tonsillar hypertrophy, it is still unclear whether SnCs play a role in these pathological conditions [182, 183]. Despite knowing that tonsillar mesenchymal stem cells can undergo cellular senescence [184, 185], the implications of cellular senescence in alterations of the function of NALT has not yet been studied.
Extensive studies in mice show that GALT exhibits a similar age-associated alteration in the cellular composition and decline in functionality like many of the other parts of the immune system. There is a decline in naïve T-cell and B-cell repertoires which are primarily replaced by memory cells [186, 187]. An age-dependent impairment in proliferative response to mitogenic stimulus is also seen in GALT [188]. There is a quantitative decline in dendritic cells accompanied by impaired functionality [189, 190] that yields a similarly impaired priming of T-cells, which is seen in the aged spleen [135, 140, 141]. This impaired immune function, with possible senescence accumulation could explain the age-associated increased rate of cancer incidence in the gastrointestinal tract.
Despite the lack of direct evidence, with the support of pre-existing knowledge of age-associated functional decline and senescence accumulation in organs [191–194] and systemic vasculature [195, 196] associated with these mucosal lymphoid tissues, it is exceedingly convincing that there is an age-dependent accumulation of SnCs in these sites and/or that their functionality is somehow impacted by this accumulation. A speculative supporting argument for this is that the mucosal surfaces are exposed to more environmental stressors than most other organs, which could possibly cause low-grade chronic activation of their immune system and SnCs accumulation. This could explain why we see a relatively early onset in the aging of the mucosal immune system compared to the systemic immune system [180, 181, 186, 197].
Apart from all the circumstantial and correlative evidence, more studies are required to further our understanding of the role of cellular senescence in age-associated changes in MALT and how or if senolytics can rejuvenate them.
Conclusion
As summarized in (Table 1), even at an organ level, the age-associated changes that contribute to immunosenescence are multifaceted with a wide variety of undesirable phenotypic manifestations. Thus, it would be ill-advised to address each of these problems individually. A more feasible and effective way to deal with immunosenescence would be to tackle the fundamental aspects of aging that drive immunosenescence. With studies showing that clearing SnCs can rejuvenate entire tissues and organs of the aged immune system [63, 140], cellular senescence is certainly one such fundamental aspect, which has the potential to address immunosenescence.
Table 1. Age-associated changes in the lymphoid organs that contribute to immunosenescence.
| Organ | Age-Associated Changes | References |
| Bone Marrow | ↑ Senescent Hematopoietic Stem Cells | [106, 198] |
| ↑ Senescent Mesenchymal Stem Cells | [95] | |
| ↑ Adiposity | [83, 88, 99] | |
| ↑ Myelopoiesis | [88] [78, 80, 82] | |
| ↓ Lymphopoiesis | [88] | |
| ↑ Oxidative Stress | [95, 96] | |
| ↑ DNA damage | [63, 94, 95, 108, 199] | |
| ↑ Inflammation | [95, 102] | |
| ↓ HSC functionality | [63, 68, 77, 200] | |
| Thymus | ↓ Structural Integrity | [111, 112] |
| ↑ Senescent Thymic Epithelial Cells | [121] | |
| ↑ Adipocytes | [112] | |
| ↑ Fibrosis | [129, 201] | |
| ↑ Inflammation | [122] | |
| ↑ DNA damage | [121] | |
| ↑ Oxidative Stress | [121] | |
| ↓ Naïve T-cell turnover | [125, 126] | |
| Spleen | ↓ Structural Integrity | [135] |
| ↓ Macrophage Phagocytosis | [139] | |
| ↑ Cellular Senescence | [50, 137] | |
| ↓ Migration of B-cells | [135, 141] | |
| ↓ Antigen Presenting Functionality | [135, 144] | |
| ↓ Recruitment of T-cells | [143] | |
| Lymph Nodes | ↓ Number | [135, 155] |
| ↓ Structural Integrity | [135, 156] | |
| ↓ Functionality | [158, 162, 202] | |
| ↑ Adiposity | [155, 156, 158] | |
| ↑ Fibrosis | [155, 156, 158] | |
| Mucosa Associated Lymphoid Tissue | ↓ Naïve B-cell repertoire | [186] |
| ↓ Naïve T-cell repertoire | [186] | |
| ↑ Memory B-cells | [186] | |
| ↑ Memory T-cells | [186] | |
| ↓ Functionality | [188, 189] | |
| ↓ Dendritic Cell Number | [190] | |
| ↓ Dendritic Cell Functionality | [189, 190] |
Cellular senescence, because of its involvement in several age-related dysfunctions and disorders, has become an essential area of interest in the field of aging research. Despite a great deal of assimilated knowledge on this phenomenon, there still remain unanswered questions. The role of cellular senescence in immunosenescence is one such key area needing further exploration. With few publications addressing the direct involvement of cellular senescence in specified immunological contexts, and many more studies providing evidence for a possible role of cellular senescence in impeding the function of the immune system, this is an area of research that deserves further exploration and an investment of resources.
In this proposed pursuit, there are several “low-hanging fruit”. A few such addressable questions include: Do SnCs play a direct or indirect role in age-related disparities seen in inflammatory pathological conditions like sepsis? Does SnCs accumulation in the peripheral tissues of the body impact the functionality of immune cells in the central nervous system? Can clearing SnCs hinder the pace of thymic involution? Can clearing SnCs in combination with thymic rejuvenation therapies in the elderly improve thymic function? Does cellular senescence drive age-associated autoimmunity? Can clearing SnCs or inhibiting SASP boost the functionality of different immune cells? Does cellular senescence play a direct role in the impaired vaccination efficacy in the elderly? Is there a senostatic/senolytic regimen that can be followed before and after vaccination to boost its efficacy in the elderly?
The increasing array of genetic models of SnC clearance along with a growing panel of senolytic and senostatic agents, provide a unique opportunity for scientists to answer these questions to lay a strong foundation to this new avenue of research in immunosenescence. Ultimately, gaining a deeper understanding of the interaction between cellular senescence and immunosenescence will help in the development of improved therapeutics that will aid in the conservation of our vitality as we age.
Author Contributions
V.B. conceived and wrote the manuscript. D.Z. conceived and revised the manuscript. T.C.F. reviewed and edited the manuscript.
Acknowledgments
We thank Sahana Manohar Sindhu, Janet Wiegand and Alexandra M. Fahnlander for their critical review of the manuscript. In addition, we thank all the reviewers of the journal for their efforts to review our manuscript.
Conflicts of Interest
D.Z. is an inventor of a pending patent application for use of Bcl-xL proteolysis targeting chimeras (PROTACs) as senolytic agents, and a co-founder and stockholder of Unity Biotechnology that develops senolytics to treat age-related diseases. The other authors declare no competing interests.
Funding
This study was supported by US National Institutes of Health (NIH) grants R01CA211963 (D.Z.), R01CA219836 (D.Z.), R01AG03801 (D.Z.), AG037984 (T.C.F.), AG052258 (T.C.F.), P30AG028740 (T.C.F.); and the Evelyn F. McKnight Brain Research Foundation.
References
- 1. Franceschi C, Garagnani P, Morsiani C, Conte M, Santoro A, Grignolio A, Monti D, Capri M, Salvioli S. The Continuum of Aging and Age-Related Diseases: Common Mechanisms but Different Rates. Front Med (Lausanne). 2018; 5:61. https://doi.org/10.3389/fmed.2018.00061 [PubMed]
- 2. United Nations, Department of Economic and Social Affairs, Population Division (2019). World Population Ageing 2019: Highlights (ST/ESA/SER.A/430). https://www.un.org/en/development/desa/population/publications/pdf/ageing/WorldPopulationAgeing2019-Highlights.pdf.
- 3. Childs BG, Durik M, Baker DJ, van Deursen JM. Cellular senescence in aging and age-related disease: from mechanisms to therapy. Nat Med. 2015; 21:1424–35. https://doi.org/10.1038/nm.4000 [PubMed]
- 4. Office of National Statistics. Overview of the UK population: January 2021. 2021. https://www.ons.gov.uk/peoplepopulationandcommunity/populationandmigration/populationestimates/articles/overviewoftheukpopulation/january2021.
- 5. Roberts AW, Bureau USC, American Community S. The population 65 years and older in the United States: 2016. https://www.census.gov/content/dam/Census/library/publications/2018/acs/ACS-38.pdf.
- 6. Vespa J, Armstrong DM, Medina L, Bureau USC. Demographic turning points for the United States: population projections for 2020 to 2060. https://www.census.gov/library/publications/2020/demo/p25-1144.html.
- 7. Kontis V, Bennett JE, Mathers CD, Li G, Foreman K, Ezzati M. Future life expectancy in 35 industrialised countries: projections with a Bayesian model ensemble. Lancet. 2017; 389:1323–35. https://doi.org/10.1016/S0140-6736(16)32381-9 [PubMed]
- 8. Piper MD, Bartke A. Diet and aging. Cell Metab. 2008; 8:99–104. https://doi.org/10.1016/j.cmet.2008.06.012 [PubMed]
- 9. Ribarič S. Diet and aging. Oxid Med Cell Longev. 2012; 2012:741468. https://doi.org/10.1155/2012/741468 [PubMed]
- 10. Garatachea N, Pareja-Galeano H, Sanchis-Gomar F, Santos-Lozano A, Fiuza-Luces C, Morán M, Emanuele E, Joyner MJ, Lucia A. Exercise attenuates the major hallmarks of aging. Rejuvenation Res. 2015; 18:57–89. https://doi.org/10.1089/rej.2014.1623 [PubMed]
- 11. Schafer MJ, White TA, Evans G, Tonne JM, Verzosa GC, Stout MB, Mazula DL, Palmer AK, Baker DJ, Jensen MD, Torbenson MS, Miller JD, Ikeda Y, et al. Exercise Prevents Diet-Induced Cellular Senescence in Adipose Tissue. Diabetes. 2016; 65:1606–15. https://doi.org/10.2337/db15-0291 [PubMed]
- 12. Davidsohn N, Pezone M, Vernet A, Graveline A, Oliver D, Slomovic S, Punthambaker S, Sun X, Liao R, Bonventre JV, Church GM. A single combination gene therapy treats multiple age-related diseases. Proc Natl Acad Sci U S A. 2019; 116:23505–11. https://doi.org/10.1073/pnas.1910073116 [PubMed]
- 13. Qian M, Liu B. Pharmaceutical Intervention of Aging. Adv Exp Med Biol. 2018; 1086:235–54. https://doi.org/10.1007/978-981-13-1117-8_15 [PubMed]
- 14. He Y, Li W, Lv D, Zhang X, Zhang X, Ortiz YT, Budamagunta V, Campisi J, Zheng G, Zhou D. Inhibition of USP7 activity selectively eliminates senescent cells in part via restoration of p53 activity. Aging Cell. 2020; 19:e13117. https://doi.org/10.1111/acel.13117 [PubMed]
- 15. He Y, Zhang X, Chang J, Kim HN, Zhang P, Wang Y, Khan S, Liu X, Zhang X, Lv D, Song L, Li W, Thummuri D, et al. Using proteolysis-targeting chimera technology to reduce navitoclax platelet toxicity and improve its senolytic activity. Nat Commun. 2020; 11:1996. https://doi.org/10.1038/s41467-020-15838-0 [PubMed]
- 16. Hickson LJ, Langhi Prata LGP, Bobart SA, Evans TK, Giorgadze N, Hashmi SK, Herrmann SM, Jensen MD, Jia Q, Jordan KL, Kellogg TA, Khosla S, Koerber DM, et al. Senolytics decrease senescent cells in humans: Preliminary report from a clinical trial of Dasatinib plus Quercetin in individuals with diabetic kidney disease. EBioMedicine. 2019; 47:446–56. https://doi.org/10.1016/j.ebiom.2019.08.069 [PubMed]
- 17. Nagata S, Hanayama R, Kawane K. Autoimmunity and the clearance of dead cells. Cell. 2010; 140:619–30. https://doi.org/10.1016/j.cell.2010.02.014 [PubMed]
- 18. Kang TW, Yevsa T, Woller N, Hoenicke L, Wuestefeld T, Dauch D, Hohmeyer A, Gereke M, Rudalska R, Potapova A, Iken M, Vucur M, Weiss S, et al. Senescence surveillance of pre-malignant hepatocytes limits liver cancer development. Nature. 2011; 479:547–51. https://doi.org/10.1038/nature10599 [PubMed]
- 19. Yevsa T, Kang TW, Zender L. Immune surveillance of pre-cancerous senescent hepatocytes limits hepatocellular carcinoma development. Oncoimmunology. 2012; 1:398–99. https://doi.org/10.4161/onci.19128 [PubMed]
- 20. Prata LGP, Ovsyannikova IG, Tchkonia T, Kirkland JL. Senescent cell clearance by the immune system: Emerging therapeutic opportunities. Semin Immunol. 2018; 40:101275. https://doi.org/10.1016/j.smim.2019.04.003 [PubMed]
- 21. Song P, An J, Zou MH. Immune Clearance of Senescent Cells to Combat Ageing and Chronic Diseases. Cells. 2020; 9:671. https://doi.org/10.3390/cells9030671 [PubMed]
- 22. Kale A, Sharma A, Stolzing A, Desprez PY, Campisi J. Role of immune cells in the removal of deleterious senescent cells. Immun Ageing. 2020; 17:16. https://doi.org/10.1186/s12979-020-00187-9 [PubMed]
- 23. Abbas AR, Baldwin D, Ma Y, Ouyang W, Gurney A, Martin F, Fong S, van Lookeren Campagne M, Godowski P, Williams PM, Chan AC, Clark HF. Immune response in silico (IRIS): immune-specific genes identified from a compendium of microarray expression data. Genes Immun. 2005; 6:319–31. https://doi.org/10.1038/sj.gene.6364173 [PubMed]
- 24. Crimmins EM. Lifespan and Healthspan: Past, Present, and Promise. Gerontologist. 2015; 55:901–11. https://doi.org/10.1093/geront/gnv130 [PubMed]
- 25. Nacionales DC, Gentile LF, Vanzant E, Lopez MC, Cuenca A, Cuenca AG, Ungaro R, Li Y, Baslanti TO, Bihorac A, Moore FA, Baker HV, Leeuwenburgh C, et al. Aged mice are unable to mount an effective myeloid response to sepsis. J Immunol. 2014; 192:612–22. https://doi.org/10.4049/jimmunol.1302109 [PubMed]
- 26. Brubaker AL, Rendon JL, Ramirez L, Choudhry MA, Kovacs EJ. Reduced neutrophil chemotaxis and infiltration contributes to delayed resolution of cutaneous wound infection with advanced age. J Immunol. 2013; 190:1746–57. https://doi.org/10.4049/jimmunol.1201213 [PubMed]
- 27. Iuliano AD, Roguski KM, Chang HH, Muscatello DJ, Palekar R, Tempia S, Cohen C, Gran JM, Schanzer D, Cowling BJ, Wu P, Kyncl J, Ang LW, et al, and Global Seasonal Influenza-associated Mortality Collaborator Network. Estimates of global seasonal influenza-associated respiratory mortality: a modelling study. Lancet. 2018; 391:1285–300. https://doi.org/10.1016/S0140-6736(17)33293-2 [PubMed]
- 28. Thompson WW, Shay DK, Weintraub E, Brammer L, Cox N, Anderson LJ, Fukuda K. Mortality associated with influenza and respiratory syncytial virus in the United States. JAMA. 2003; 289:179–86. https://doi.org/10.1001/jama.289.2.179 [PubMed]
- 29. Demicheli V, Jefferson T, Di Pietrantonj C, Ferroni E, Thorning S, Thomas RE, Rivetti A. Vaccines for preventing influenza in the elderly. Cochrane Database Syst Rev. 2018; 2:CD004876. [PubMed]
- 30. Goodwin K, Viboud C, Simonsen L. Antibody response to influenza vaccination in the elderly: a quantitative review. Vaccine. 2006; 24:1159–69. https://doi.org/10.1016/j.vaccine.2005.08.105 [PubMed]
- 31. Yang X, Yu Y, Xu J, Shu H, Xia J, Liu H, Wu Y, Zhang L, Yu Z, Fang M, Yu T, Wang Y, Pan S, et al. Clinical course and outcomes of critically ill patients with SARS-CoV-2 pneumonia in Wuhan, China: a single-centered, retrospective, observational study. Lancet Respir Med. 2020; 8:475–81. https://doi.org/10.1016/S2213-2600(20)30079-5 [PubMed]
- 32. Ovadya Y, Landsberger T, Leins H, Vadai E, Gal H, Biran A, Yosef R, Sagiv A, Agrawal A, Shapira A, Windheim J, Tsoory M, Schirmbeck R, et al. Impaired immune surveillance accelerates accumulation of senescent cells and aging. Nat Commun. 2018; 9:5435. https://doi.org/10.1038/s41467-018-07825-3 [PubMed]
- 33. Pera A, Campos C, López N, Hassouneh F, Alonso C, Tarazona R, Solana R. Immunosenescence: Implications for response to infection and vaccination in older people. Maturitas. 2015; 82:50–55. https://doi.org/10.1016/j.maturitas.2015.05.004 [PubMed]
- 34. Karin O, Agrawal A, Porat Z, Krizhanovsky V, Alon U. Senescent cell turnover slows with age providing an explanation for the Gompertz law. Nat Commun. 2019; 10:5495. https://doi.org/10.1038/s41467-019-13192-4 [PubMed]
- 35. Keilich SR, Bartley JM, Haynes L. Diminished immune responses with aging predispose older adults to common and uncommon influenza complications. Cell Immunol. 2019; 345:103992. https://doi.org/10.1016/j.cellimm.2019.103992 [PubMed]
- 36. Qin C, Zhou L, Hu Z, Zhang S, Yang S, Tao Y, Xie C, Ma K, Shang K, Wang W, Tian DS. Dysregulation of Immune Response in Patients With Coronavirus 2019 (COVID-19) in Wuhan, China. Clin Infect Dis. 2020; 71:762–68. https://doi.org/10.1093/cid/ciaa248 [PubMed]
- 37. Goronzy JJ, Weyand CM. Immune aging and autoimmunity. Cell Mol Life Sci. 2012; 69:1615–23. [PubMed]
- 38. Goronzy JJ, Shao L, Weyand CM. Immune aging and rheumatoid arthritis. Rheum Dis Clin North Am. 2010; 36:297–310. https://doi.org/10.1016/j.rdc.2010.03.001 [PubMed]
- 39. Sidler C, Wóycicki R, Ilnytskyy Y, Metz G, Kovalchuk I, Kovalchuk O. Immunosenescence is associated with altered gene expression and epigenetic regulation in primary and secondary immune organs. Front Genet. 2013; 4:211. https://doi.org/10.3389/fgene.2013.00211 [PubMed]
- 40. Turner JE. Is immunosenescence influenced by our lifetime "dose" of exercise? Biogerontology. 2016; 17:581–602. [PubMed]
- 41. Maijó M, Clements SJ, Ivory K, Nicoletti C, Carding SR. Nutrition, diet and immunosenescence. Mech Ageing Dev. 2014; 136–137:116–28. https://doi.org/10.1016/j.mad.2013.12.003 [PubMed]
- 42. Caruso C, Accardi G, Virruso C, Candore G. Sex, gender and immunosenescence: a key to understand the different lifespan between men and women? Immun Ageing. 2013; 10:20. https://doi.org/10.1186/1742-4933-10-20 [PubMed]
- 43. Hayflick L, Moorhead PS. The serial cultivation of human diploid cell strains. Exp Cell Res. 1961; 25:585–621. https://doi.org/10.1016/0014-4827(61)90192-6 [PubMed]
- 44. Campisi J, d'Adda di Fagagna F. Cellular senescence: when bad things happen to good cells. Nat Rev Mol Cell Biol. 2007; 8:729–40. https://doi.org/10.1038/nrm2233 [PubMed]
- 45. Collado M, Blasco MA, Serrano M. Cellular senescence in cancer and aging. Cell. 2007; 130:223–33. https://doi.org/10.1016/j.cell.2007.07.003 [PubMed]
- 46. Krizhanovsky V, Yon M, Dickins RA, Hearn S, Simon J, Miething C, Yee H, Zender L, Lowe SW. Senescence of activated stellate cells limits liver fibrosis. Cell. 2008; 134:657–67. https://doi.org/10.1016/j.cell.2008.06.049 [PubMed]
- 47. Kim KH, Chen CC, Monzon RI, Lau LF. Matricellular protein CCN1 promotes regression of liver fibrosis through induction of cellular senescence in hepatic myofibroblasts. Mol Cell Biol. 2013; 33:2078–90. https://doi.org/10.1128/MCB.00049-13 [PubMed]
- 48. Cui S, Xue L, Yang F, Dai S, Han Z, Liu K, Liu B, Yuan Q, Cui Z, Zhang Y, Xu F, Chen Y. Postinfarction Hearts Are Protected by Premature Senescent Cardiomyocytes Via GATA 4-Dependent CCN 1 Secretion. J Am Heart Assoc. 2018; 7:e009111. https://doi.org/10.1161/JAHA.118.009111 [PubMed]
- 49. Demaria M, Ohtani N, Youssef SA, Rodier F, Toussaint W, Mitchell JR, Laberge RM, Vijg J, Van Steeg H, Dollé ME, Hoeijmakers JH, de Bruin A, Hara E, Campisi J. An essential role for senescent cells in optimal wound healing through secretion of PDGF-AA. Dev Cell. 2014; 31:722–33. https://doi.org/10.1016/j.devcel.2014.11.012 [PubMed]
- 50. Baker DJ, Childs BG, Durik M, Wijers ME, Sieben CJ, Zhong J, Saltness RA, Jeganathan KB, Verzosa GC, Pezeshki A, Khazaie K, Miller JD, van Deursen JM. Naturally occurring p16(Ink4a)-positive cells shorten healthy lifespan. Nature. 2016; 530:184–89. https://doi.org/10.1038/nature16932 [PubMed]
- 51. Coppé JP, Desprez PY, Krtolica A, Campisi J. The senescence-associated secretory phenotype: the dark side of tumor suppression. Annu Rev Pathol. 2010; 5:99–118. https://doi.org/10.1146/annurev-pathol-121808-102144 [PubMed]
- 52. Lasry A, Ben-Neriah Y. Senescence-associated inflammatory responses: aging and cancer perspectives. Trends Immunol. 2015; 36:217–28. https://doi.org/10.1016/j.it.2015.02.009 [PubMed]
- 53. He S, Sharpless NE. Senescence in Health and Disease. Cell. 2017; 169:1000–11. https://doi.org/10.1016/j.cell.2017.05.015 [PubMed]
- 54. Lewis-McDougall FC, Ruchaya PJ, Domenjo-Vila E, Shin Teoh T, Prata L, Cottle BJ, Clark JE, Punjabi PP, Awad W, Torella D, Tchkonia T, Kirkland JL, Ellison-Hughes GM. Aged-senescent cells contribute to impaired heart regeneration. Aging Cell. 2019; 18:e12931. https://doi.org/10.1111/acel.12931 [PubMed]
- 55. Ogrodnik M, Zhu Y, Langhi LGP, Tchkonia T, Krüger P, Fielder E, Victorelli S, Ruswhandi RA, Giorgadze N, Pirtskhalava T, Podgorni O, Enikolopov G, Johnson KO, et al. Obesity-Induced Cellular Senescence Drives Anxiety and Impairs Neurogenesis. Cell Metab. 2019; 29:1233. https://doi.org/10.1016/j.cmet.2019.01.013 [PubMed]
- 56. Minamino T, Miyauchi H, Yoshida T, Ishida Y, Yoshida H, Komuro I. Endothelial cell senescence in human atherosclerosis: role of telomere in endothelial dysfunction. Circulation. 2002; 105:1541–44. https://doi.org/10.1161/01.cir.0000013836.85741.17 [PubMed]
- 57. Chimenti C, Kajstura J, Torella D, Urbanek K, Heleniak H, Colussi C, Di Meglio F, Nadal-Ginard B, Frustaci A, Leri A, Maseri A, Anversa P. Senescence and death of primitive cells and myocytes lead to premature cardiac aging and heart failure. Circ Res. 2003; 93:604–13. https://doi.org/10.1161/01.RES.0000093985.76901.AF [PubMed]
- 58. Xu M, Pirtskhalava T, Farr JN, Weigand BM, Palmer AK, Weivoda MM, Inman CL, Ogrodnik MB, Hachfeld CM, Fraser DG, Onken JL, Johnson KO, Verzosa GC, et al. Senolytics improve physical function and increase lifespan in old age. Nat Med. 2018; 24:1246–56. https://doi.org/10.1038/s41591-018-0092-9 [PubMed]
- 59. Sun R, Zhu B, Xiong K, Sun Y, Shi D, Chen L, Zhang Y, Li Z, Xue L. Senescence as a novel mechanism involved in β-adrenergic receptor mediated cardiac hypertrophy. PLoS One. 2017; 12:e0182668. https://doi.org/10.1371/journal.pone.0182668 [PubMed]
- 60. Nelson G, Wordsworth J, Wang C, Jurk D, Lawless C, Martin-Ruiz C, von Zglinicki T. A senescent cell bystander effect: senescence-induced senescence. Aging Cell. 2012; 11:345–49. https://doi.org/10.1111/j.1474-9726.2012.00795.x [PubMed]
- 61. da Silva PFL, Ogrodnik M, Kucheryavenko O, Glibert J, Miwa S, Cameron K, Ishaq A, Saretzki G, Nagaraja-Grellscheid S, Nelson G, von Zglinicki T. The bystander effect contributes to the accumulation of senescent cells in vivo. Aging Cell. 2019; 18:e12848. https://doi.org/10.1111/acel.12848 [PubMed]
- 62. Peng X, Wu Y, Brouwer U, van Vliet T, Wang B, Demaria M, Barazzuol L, Coppes RP. Cellular senescence contributes to radiation-induced hyposalivation by affecting the stem/progenitor cell niche. Cell Death Dis. 2020; 11:854. https://doi.org/10.1038/s41419-020-03074-9 [PubMed]
- 63. Chang J, Wang Y, Shao L, Laberge RM, Demaria M, Campisi J, Janakiraman K, Sharpless NE, Ding S, Feng W, Luo Y, Wang X, Aykin-Burns N, et al. Clearance of senescent cells by ABT263 rejuvenates aged hematopoietic stem cells in mice. Nat Med. 2016; 22:78–83. https://doi.org/10.1038/nm.4010 [PubMed]
- 64. Sagiv A, Burton DG, Moshayev Z, Vadai E, Wensveen F, Ben-Dor S, Golani O, Polic B, Krizhanovsky V. NKG2D ligands mediate immunosurveillance of senescent cells. Aging (Albany NY). 2016; 8:328–44. https://doi.org/10.18632/aging.100897 [PubMed]
- 65. Fuentes E, Fuentes M, Alarcón M, Palomo I. Immune System Dysfunction in the Elderly. An Acad Bras Cienc. 2017; 89:285–99. https://doi.org/10.1590/0001-3765201720160487 [PubMed]
- 66. Pereira BI, Devine OP, Vukmanovic-Stejic M, Chambers ES, Subramanian P, Patel N, Virasami A, Sebire NJ, Kinsler V, Valdovinos A, LeSaux CJ, Passos JF, Antoniou A, et al. Senescent cells evade immune clearance via HLA-E-mediated NK and CD8+ T cell inhibition. Nat Commun. 2019; 10:2387. https://doi.org/10.1038/s41467-019-10335-5 [PubMed]
- 67. Muñoz DP, Yannone SM, Daemen A, Sun Y, Vakar-Lopez F, Kawahara M, Freund AM, Rodier F, Wu JD, Desprez PY, Raulet DH, Nelson PS, van 't Veer LJ, et al. Targetable mechanisms driving immunoevasion of persistent senescent cells link chemotherapy-resistant cancer to aging. JCI Insight. 2019; 5:e124716. https://doi.org/10.1172/jci.insight.124716 [PubMed]
- 68. Janzen V, Forkert R, Fleming HE, Saito Y, Waring MT, Dombkowski DM, Cheng T, DePinho RA, Sharpless NE, Scadden DT. Stem-cell ageing modified by the cyclin-dependent kinase inhibitor p16INK4a. Nature. 2006; 443:421–26. https://doi.org/10.1038/nature05159 [PubMed]
- 69. Molofsky AV, Slutsky SG, Joseph NM, He S, Pardal R, Krishnamurthy J, Sharpless NE, Morrison SJ. Increasing p16INK4a expression decreases forebrain progenitors and neurogenesis during ageing. Nature. 2006; 443:448–52. https://doi.org/10.1038/nature05091 [PubMed]
- 70. Josephson AM, Bradaschia-Correa V, Lee S, Leclerc K, Patel KS, Muinos Lopez E, Litwa HP, Neibart SS, Kadiyala M, Wong MZ, Mizrahi MM, Yim NL, Ramme AJ, et al. Age-related inflammation triggers skeletal stem/progenitor cell dysfunction. Proc Natl Acad Sci U S A. 2019; 116:6995–7004. https://doi.org/10.1073/pnas.1810692116 [PubMed]
- 71. Zhao J, Zhang L, Lu A, Han Y, Colangelo D, Bukata C, Scibetta A, Yousefzadeh MJ, Li X, Gurkar AU, McGowan SJ, Angelini L, O'Kelly R, et al. ATM is a key driver of NF-κB-dependent DNA-damage-induced senescence, stem cell dysfunction and aging. Aging (Albany NY). 2020; 12:4688–710. https://doi.org/10.18632/aging.102863 [PubMed]
- 72. Krishnamurthy J, Ramsey MR, Ligon KL, Torrice C, Koh A, Bonner-Weir S, Sharpless NE. p16INK4a induces an age-dependent decline in islet regenerative potential. Nature. 2006; 443:453–57. https://doi.org/10.1038/nature05092 [PubMed]
- 73. Zhao E, Xu H, Wang L, Kryczek I, Wu K, Hu Y, Wang G, Zou W. Bone marrow and the control of immunity. Cell Mol Immunol. 2012; 9:11–19. https://doi.org/10.1038/cmi.2011.47 [PubMed]
- 74. Muschler GF, Nitto H, Boehm CA, Easley KA. Age- and gender-related changes in the cellularity of human bone marrow and the prevalence of osteoblastic progenitors. J Orthop Res. 2001; 19:117–25. https://doi.org/10.1016/S0736-0266(00)00010-3 [PubMed]
- 75. Pritz T, Weinberger B, Grubeck-Loebenstein B. The aging bone marrow and its impact on immune responses in old age. Immunol Lett. 2014; 162:310–15. https://doi.org/10.1016/j.imlet.2014.06.016 [PubMed]
- 76. Hoffman CM, Han J, Calvi LM. Impact of aging on bone, marrow and their interactions. Bone. 2019; 119:1–7. https://doi.org/10.1016/j.bone.2018.07.012 [PubMed]
- 77. Rossi DJ, Bryder D, Zahn JM, Ahlenius H, Sonu R, Wagers AJ, Weissman IL. Cell intrinsic alterations underlie hematopoietic stem cell aging. Proc Natl Acad Sci U S A. 2005; 102:9194–99. https://doi.org/10.1073/pnas.0503280102 [PubMed]
- 78. Pang WW, Price EA, Sahoo D, Beerman I, Maloney WJ, Rossi DJ, Schrier SL, Weissman IL. Human bone marrow hematopoietic stem cells are increased in frequency and myeloid-biased with age. Proc Natl Acad Sci U S A. 2011; 108:20012–17. https://doi.org/10.1073/pnas.1116110108 [PubMed]
- 79. Ogawa T, Kitagawa M, Hirokawa K. Age-related changes of human bone marrow: a histometric estimation of proliferative cells, apoptotic cells, T cells, B cells and macrophages. Mech Ageing Dev. 2000; 117:57–68. https://doi.org/10.1016/s0047-6374(00)00137-8 [PubMed]
- 80. Liang Y, Van Zant G, Szilvassy SJ. Effects of aging on the homing and engraftment of murine hematopoietic stem and progenitor cells. Blood. 2005; 106:1479–87. https://doi.org/10.1182/blood-2004-11-4282 [PubMed]
- 81. Vas V, Senger K, Dörr K, Niebel A, Geiger H. Aging of the microenvironment influences clonality in hematopoiesis. PLoS One. 2012; 7:e42080. https://doi.org/10.1371/journal.pone.0042080 [PubMed]
- 82. Ergen AV, Boles NC, Goodell MA. Rantes/Ccl5 influences hematopoietic stem cell subtypes and causes myeloid skewing. Blood. 2012; 119:2500–09. https://doi.org/10.1182/blood-2011-11-391730 [PubMed]
- 83. Takeshita S, Fumoto T, Naoe Y, Ikeda K. Age-related marrow adipogenesis is linked to increased expression of RANKL. J Biol Chem. 2014; 289:16699–710. https://doi.org/10.1074/jbc.M114.547919 [PubMed]
- 84. Umemoto Y, Tsuji K, Yang FC, Ebihara Y, Kaneko A, Furukawa S, Nakahata T. Leptin stimulates the proliferation of murine myelocytic and primitive hematopoietic progenitor cells. Blood. 1997; 90:3438–43. [PubMed]
- 85. Shimozato T, Kincade PW. Prostaglandin E(2) and stem cell factor can deliver opposing signals to B lymphocyte precursors. Cell Immunol. 1999; 198:21–29. https://doi.org/10.1006/cimm.1999.1575 [PubMed]
- 86. Yokota T, Meka CS, Kouro T, Medina KL, Igarashi H, Takahashi M, Oritani K, Funahashi T, Tomiyama Y, Matsuzawa Y, Kincade PW. Adiponectin, a fat cell product, influences the earliest lymphocyte precursors in bone marrow cultures by activation of the cyclooxygenase-prostaglandin pathway in stromal cells. J Immunol. 2003; 171:5091–99. https://doi.org/10.4049/jimmunol.171.10.5091 [PubMed]
- 87. Poloni A, Maurizi G, Serrani F, Mancini S, Zingaretti MC, Frontini A, Cinti S, Olivieri A, Leoni P. Molecular and functional characterization of human bone marrow adipocytes. Exp Hematol. 2013; 41:558–66.e2. https://doi.org/10.1016/j.exphem.2013.02.005 [PubMed]
- 88. Wang H, Leng Y, Gong Y. Bone Marrow Fat and Hematopoiesis. Front Endocrinol (Lausanne). 2018; 9:694. https://doi.org/10.3389/fendo.2018.00694 [PubMed]
- 89. Farr JN, Fraser DG, Wang H, Jaehn K, Ogrodnik MB, Weivoda MM, Drake MT, Tchkonia T, LeBrasseur NK, Kirkland JL, Bonewald LF, Pignolo RJ, Monroe DG, Khosla S. Identification of Senescent Cells in the Bone Microenvironment. J Bone Miner Res. 2016; 31:1920–29. https://doi.org/10.1002/jbmr.2892 [PubMed]
- 90. Liu J, Ding Y, Liu Z, Liang X. Senescence in Mesenchymal Stem Cells: Functional Alterations, Molecular Mechanisms, and Rejuvenation Strategies. Front Cell Dev Biol. 2020; 8:258. https://doi.org/10.3389/fcell.2020.00258 [PubMed]
- 91. Kim HN, Xiong J, MacLeod RS, Iyer S, Fujiwara Y, Cawley KM, Han L, He Y, Thostenson JD, Ferreira E, Jilka RL, Zhou D, Almeida M, O'Brien CA. Osteocyte RANKL is required for cortical bone loss with age and is induced by senescence. JCI Insight. 2020; 5:e138815. https://doi.org/10.1172/jci.insight.138815 [PubMed]
- 92. Eyman D, Damodarasamy M, Plymate SR, Reed MJ. CCL5 secreted by senescent aged fibroblasts induces proliferation of prostate epithelial cells and expression of genes that modulate angiogenesis. J Cell Physiol. 2009; 220:376–81. https://doi.org/10.1002/jcp.21776 [PubMed]
- 93. Liu ZJ, Tan Y, Beecham GW, Seo DM, Tian R, Li Y, Vazquez-Padron RI, Pericak-Vance M, Vance JM, Goldschmidt-Clermont PJ, Livingstone AS, Velazquez OC. Notch activation induces endothelial cell senescence and pro-inflammatory response: implication of Notch signaling in atherosclerosis. Atherosclerosis. 2012; 225:296–303. https://doi.org/10.1016/j.atherosclerosis.2012.04.010 [PubMed]
- 94. Minieri V, Saviozzi S, Gambarotta G, Lo Iacono M, Accomasso L, Cibrario Rocchietti E, Gallina C, Turinetto V, Giachino C. Persistent DNA damage-induced premature senescence alters the functional features of human bone marrow mesenchymal stem cells. J Cell Mol Med. 2015; 19:734–43. https://doi.org/10.1111/jcmm.12387 [PubMed]
- 95. Gnani D, Crippa S, Della Volpe L, Rossella V, Conti A, Lettera E, Rivis S, Ometti M, Fraschini G, Bernardo ME, Di Micco R. An early-senescence state in aged mesenchymal stromal cells contributes to hematopoietic stem and progenitor cell clonogenic impairment through the activation of a pro-inflammatory program. Aging Cell. 2019; 18:e12933. https://doi.org/10.1111/acel.12933 [PubMed]
- 96. Khatri R, Krishnan S, Roy S, Chattopadhyay S, Kumar V, Mukhopadhyay A. Reactive Oxygen Species Limit the Ability of Bone Marrow Stromal Cells to Support Hematopoietic Reconstitution in Aging Mice. Stem Cells Dev. 2016; 25:948–58. https://doi.org/10.1089/scd.2015.0391 [PubMed]
- 97. Tchkonia T, Morbeck DE, Von Zglinicki T, Van Deursen J, Lustgarten J, Scrable H, Khosla S, Jensen MD, Kirkland JL. Fat tissue, aging, and cellular senescence. Aging Cell. 2010; 9:667–84. https://doi.org/10.1111/j.1474-9726.2010.00608.x [PubMed]
- 98. Farr JN, Xu M, Weivoda MM, Monroe DG, Fraser DG, Onken JL, Negley BA, Sfeir JG, Ogrodnik MB, Hachfeld CM, LeBrasseur NK, Drake MT, Pignolo RJ, et al. Corrigendum: Targeting cellular senescence prevents age-related bone loss in mice. Nat Med. 2017; 23:1384. https://doi.org/10.1038/nm1117-1384c [PubMed]
- 99. Ambrosi TH, Scialdone A, Graja A, Gohlke S, Jank AM, Bocian C, Woelk L, Fan H, Logan DW, Schürmann A, Saraiva LR, Schulz TJ. Adipocyte Accumulation in the Bone Marrow during Obesity and Aging Impairs Stem Cell-Based Hematopoietic and Bone Regeneration. Cell Stem Cell. 2017; 20:771–84.e6. https://doi.org/10.1016/j.stem.2017.02.009 [PubMed]
- 100. Naveiras O, Nardi V, Wenzel PL, Hauschka PV, Fahey F, Daley GQ. Bone-marrow adipocytes as negative regulators of the haematopoietic microenvironment. Nature. 2009; 460:259–63. https://doi.org/10.1038/nature08099 [PubMed]
- 101. Stephan RP, Reilly CR, Witte PL. Impaired ability of bone marrow stromal cells to support B-lymphopoiesis with age. Blood. 1998; 91:75–88. [PubMed]
- 102. Ueda Y, Yang K, Foster SJ, Kondo M, Kelsoe G. Inflammation controls B lymphopoiesis by regulating chemokine CXCL12 expression. J Exp Med. 2004; 199:47–58. https://doi.org/10.1084/jem.20031104 [PubMed]
- 103. Hagen M, Derudder E. Inflammation and the Alteration of B-Cell Physiology in Aging. Gerontology. 2020; 66:105–13. https://doi.org/10.1159/000501963 [PubMed]
- 104. Tang J, Nuccie BL, Ritterman I, Liesveld JL, Abboud CN, Ryan DH. TGF-beta down-regulates stromal IL-7 secretion and inhibits proliferation of human B cell precursors. J Immunol. 1997; 159:117–25. [PubMed]
- 105. Day RB, Bhattacharya D, Nagasawa T, Link DC. Granulocyte colony-stimulating factor reprograms bone marrow stromal cells to actively suppress B lymphopoiesis in mice. Blood. 2015; 125:3114–17. https://doi.org/10.1182/blood-2015-02-629444 [PubMed]
- 106. Fali T, Fabre-Mersseman V, Yamamoto T, Bayard C, Papagno L, Fastenackels S, Zoorab R, Koup RA, Boddaert J, Sauce D, Appay V. Elderly human hematopoietic progenitor cells express cellular senescence markers and are more susceptible to pyroptosis. JCI Insight. 2018; 3:e95319. https://doi.org/10.1172/jci.insight.95319 [PubMed]
- 107. Nelson G, Kucheryavenko O, Wordsworth J, von Zglinicki T. The senescent bystander effect is caused by ROS-activated NF-κB signalling. Mech Ageing Dev. 2018; 170:30–36. https://doi.org/10.1016/j.mad.2017.08.005 [PubMed]
- 108. Beerman I, Seita J, Inlay MA, Weissman IL, Rossi DJ. Quiescent hematopoietic stem cells accumulate DNA damage during aging that is repaired upon entry into cell cycle. Cell Stem Cell. 2014; 15:37–50. https://doi.org/10.1016/j.stem.2014.04.016 [PubMed]
- 109. Mohrin M, Bourke E, Alexander D, Warr MR, Barry-Holson K, Le Beau MM, Morrison CG, Passegué E. Hematopoietic stem cell quiescence promotes error-prone DNA repair and mutagenesis. Cell Stem Cell. 2010; 7:174–85. https://doi.org/10.1016/j.stem.2010.06.014 [PubMed]
- 110. Rossi DJ, Bryder D, Seita J, Nussenzweig A, Hoeijmakers J, Weissman IL. Deficiencies in DNA damage repair limit the function of haematopoietic stem cells with age. Nature. 2007; 447:725–29. https://doi.org/10.1038/nature05862 [PubMed]
- 111. Aw D, Silva AB, Maddick M, von Zglinicki T, Palmer DB. Architectural changes in the thymus of aging mice. Aging Cell. 2008; 7:158–67. https://doi.org/10.1111/j.1474-9726.2007.00365.x [PubMed]
- 112. Cavallotti C, D'Andrea V, Tonnarini G, Cavallotti C, Bruzzone P. Age-related changes in the human thymus studied with scanning electron microscopy. Microsc Res Tech. 2008; 71:573–78. https://doi.org/10.1002/jemt.20588 [PubMed]
- 113. Varecza Z, Kvell K, Talabér G, Miskei G, Csongei V, Bartis D, Anderson G, Jenkinson EJ, Pongracz JE. Multiple suppression pathways of canonical Wnt signalling control thymic epithelial senescence. Mech Ageing Dev. 2011; 132:249–56. https://doi.org/10.1016/j.mad.2011.04.007 [PubMed]
- 114. Kvell K, Varecza Z, Bartis D, Hesse S, Parnell S, Anderson G, Jenkinson EJ, Pongracz JE. Wnt4 and LAP2alpha as pacemakers of thymic epithelial senescence. PLoS One. 2010; 5:e10701. https://doi.org/10.1371/journal.pone.0010701 [PubMed]
- 115. Aw D, Taylor-Brown F, Cooper K, Palmer DB. Phenotypical and morphological changes in the thymic microenvironment from ageing mice. Biogerontology. 2009; 10:311–22. https://doi.org/10.1007/s10522-008-9182-2 [PubMed]
- 116. Shanley DP, Aw D, Manley NR, Palmer DB. An evolutionary perspective on the mechanisms of immunosenescence. Trends Immunol. 2009; 30:374–81. https://doi.org/10.1016/j.it.2009.05.001 [PubMed]
- 117. Petrie HT. Role of thymic organ structure and stromal composition in steady-state postnatal T-cell production. Immunol Rev. 2002; 189:8–19. https://doi.org/10.1034/j.1600-065x.2002.18902.x [PubMed]
- 118. Goronzy JJ, Lee WW, Weyand CM. Aging and T-cell diversity. Exp Gerontol. 2007; 42:400–06. https://doi.org/10.1016/j.exger.2006.11.016 [PubMed]
- 119. Coder BD, Wang H, Ruan L, Su DM. Thymic involution perturbs negative selection leading to autoreactive T cells that induce chronic inflammation. J Immunol. 2015; 194:5825–37. https://doi.org/10.4049/jimmunol.1500082 [PubMed]
- 120. Thomas R, Wang W, Su DM. Contributions of Age-Related Thymic Involution to Immunosenescence and Inflammaging. Immun Ageing. 2020; 17:2. https://doi.org/10.1186/s12979-020-0173-8 [PubMed]
- 121. Barbouti A, Evangelou K, Pateras IS, Papoudou-Bai A, Patereli A, Stefanaki K, Rontogianni D, Muñoz-Espín D, Kanavaros P, Gorgoulis VG. In situ evidence of cellular senescence in Thymic Epithelial Cells (TECs) during human thymic involution. Mech Ageing Dev. 2019; 177:88–90. https://doi.org/10.1016/j.mad.2018.02.005 [PubMed]
- 122. Sempowski GD, Hale LP, Sundy JS, Massey JM, Koup RA, Douek DC, Patel DD, Haynes BF. Leukemia inhibitory factor, oncostatin M, IL-6, and stem cell factor mRNA expression in human thymus increases with age and is associated with thymic atrophy. J Immunol. 2000; 164:2180–87. https://doi.org/10.4049/jimmunol.164.4.2180 [PubMed]
- 123. Griffith AV, Venables T, Shi J, Farr A, van Remmen H, Szweda L, Fallahi M, Rabinovitch P, Petrie HT. Metabolic Damage and Premature Thymus Aging Caused by Stromal Catalase Deficiency. Cell Rep. 2015; 12:1071–79. https://doi.org/10.1016/j.celrep.2015.07.008 [PubMed]
- 124. Takaba H, Takayanagi H. The Mechanisms of T Cell Selection in the Thymus. Trends Immunol. 2017; 38:805–16. https://doi.org/10.1016/j.it.2017.07.010 [PubMed]
- 125. Gui J, Zhu X, Dohkan J, Cheng L, Barnes PF, Su DM. The aged thymus shows normal recruitment of lymphohematopoietic progenitors but has defects in thymic epithelial cells. Int Immunol. 2007; 19:1201–11. https://doi.org/10.1093/intimm/dxm095 [PubMed]
- 126. Zhu X, Gui J, Dohkan J, Cheng L, Barnes PF, Su DM. Lymphohematopoietic progenitors do not have a synchronized defect with age-related thymic involution. Aging Cell. 2007; 6:663–72. https://doi.org/10.1111/j.1474-9726.2007.00325.x [PubMed]
- 127. Becklund BR, Purton JF, Ramsey C, Favre S, Vogt TK, Martin CE, Spasova DS, Sarkisyan G, LeRoy E, Tan JT, Wahlus H, Bondi-Boyd B, Luther SA, Surh CD. The aged lymphoid tissue environment fails to support naïve T cell homeostasis. Sci Rep. 2016; 6:30842. https://doi.org/10.1038/srep30842 [PubMed]
- 128. Mackall CL, Punt JA, Morgan P, Farr AG, Gress RE. Thymic function in young/old chimeras: substantial thymic T cell regenerative capacity despite irreversible age-associated thymic involution. Eur J Immunol. 1998; 28:1886–93. https://doi.org/10.1002/(SICI)1521-4141(199806)28:06<1886::AID-IMMU1886>3.0.CO;2-M [PubMed]
- 129. Thompson HL, Smithey MJ, Uhrlaub JL, Jeftić I, Jergović M, White SE, Currier N, Lang AM, Okoye A, Park B, Picker LJ, Surh CD, Nikolich-Žugich J. Lymph nodes as barriers to T-cell rejuvenation in aging mice and nonhuman primates. Aging Cell. 2019; 18:e12865. https://doi.org/10.1111/acel.12865 [PubMed]
- 130. Fontana L, Nehme J, Demaria M. Caloric restriction and cellular senescence. Mech Ageing Dev. 2018; 176:19–23. https://doi.org/10.1016/j.mad.2018.10.005 [PubMed]
- 131. Longo VD, Cortellino S. Fasting, dietary restriction, and immunosenescence. J Allergy Clin Immunol. 2020; 146:1002–04. https://doi.org/10.1016/j.jaci.2020.07.035 [PubMed]
- 132. Chung HY, Kim DH, Bang E, Yu BP. Impacts of Calorie Restriction and Intermittent Fasting on Health and Diseases: Current Trends. Nutrients. 2020; 12:2948. https://doi.org/10.3390/nu12102948 [PubMed]
- 133. Yang H, Youm YH, Dixit VD. Inhibition of thymic adipogenesis by caloric restriction is coupled with reduction in age-related thymic involution. J Immunol. 2009; 183:3040–52. https://doi.org/10.4049/jimmunol.0900562 [PubMed]
- 134. Bronte V, Pittet MJ. The spleen in local and systemic regulation of immunity. Immunity. 2013; 39:806–18. https://doi.org/10.1016/j.immuni.2013.10.010 [PubMed]
- 135. Turner VM, Mabbott NA. Influence of ageing on the microarchitecture of the spleen and lymph nodes. Biogerontology. 2017; 18:723–38. https://doi.org/10.1007/s10522-017-9707-7 [PubMed]
- 136. Aw D, Hilliard L, Nishikawa Y, Cadman ET, Lawrence RA, Palmer DB. Disorganization of the splenic microanatomy in ageing mice. Immunology. 2016; 148:92–101. https://doi.org/10.1111/imm.12590 [PubMed]
- 137. Wang C, Jurk D, Maddick M, Nelson G, Martin-Ruiz C, von Zglinicki T. DNA damage response and cellular senescence in tissues of aging mice. Aging Cell. 2009; 8:311–23. https://doi.org/10.1111/j.1474-9726.2009.00481.x [PubMed]
- 138. Park J, Miyakawa T, Shiokawa A, Nakajima-Adachi H, Tanokura M, Hachimura S. Splenic stromal cells from aged mice produce higher levels of IL-6 compared to young mice. Mediators Inflamm. 2014; 2014:826987. https://doi.org/10.1155/2014/826987 [PubMed]
- 139. Birjandi SZ, Ippolito JA, Ramadorai AK, Witte PL. Alterations in marginal zone macrophages and marginal zone B cells in old mice. J Immunol. 2011; 186:3441–51. https://doi.org/10.4049/jimmunol.1001271 [PubMed]
- 140. Palacio L, Goyer ML, Maggiorani D, Espinosa A, Villeneuve N, Bourbonnais S, Moquin-Beaudry G, Le O, Demaria M, Davalos AR, Decaluwe H, Beauséjour C. Restored immune cell functions upon clearance of senescence in the irradiated splenic environment. Aging Cell. 2019; 18:e12971. https://doi.org/10.1111/acel.12971 [PubMed]
- 141. Turner VM, Mabbott NA. Ageing adversely affects the migration and function of marginal zone B cells. Immunology. 2017; 151:349–62. https://doi.org/10.1111/imm.12737 [PubMed]
- 142. Turner DL, Bickham KL, Farber DL, Lefrançois L. Splenic priming of virus-specific CD8 T cells following influenza virus infection. J Virol. 2013; 87:4496–506. https://doi.org/10.1128/JVI.03413-12 [PubMed]
- 143. Lefebvre JS, Maue AC, Eaton SM, Lanthier PA, Tighe M, Haynes L. The aged microenvironment contributes to the age-related functional defects of CD4 T cells in mice. Aging Cell. 2012; 11:732–40. https://doi.org/10.1111/j.1474-9726.2012.00836.x [PubMed]
- 144. Li G, Smithey MJ, Rudd BD, Nikolich-Žugich J. Age-associated alterations in CD8α+ dendritic cells impair CD8 T-cell expansion in response to an intracellular bacterium. Aging Cell. 2012; 11:968–77. https://doi.org/10.1111/j.1474-9726.2012.00867.x [PubMed]
- 145. Zhu Y, Tchkonia T, Pirtskhalava T, Gower AC, Ding H, Giorgadze N, Palmer AK, Ikeno Y, Hubbard GB, Lenburg M, O'Hara SP, LaRusso NF, Miller JD, et al. The Achilles’ heel of senescent cells: from transcriptome to senolytic drugs. Aging Cell. 2015; 14:644–58. https://doi.org/10.1111/acel.12344 [PubMed]
- 146. Zhu Y, Doornebal EJ, Pirtskhalava T, Giorgadze N, Wentworth M, Fuhrmann-Stroissnigg H, Niedernhofer LJ, Robbins PD, Tchkonia T, Kirkland JL. New agents that target senescent cells: the flavone, fisetin, and the BCL-XL inhibitors, A1331852 and A1155463. Aging (Albany NY). 2017; 9:955–63. https://doi.org/10.18632/aging.101202 [PubMed]
- 147. Martin N, Soriani O, Bernard D. Cardiac Glycosides as Senolytic Compounds. Trends Mol Med. 2020; 26:243–45. https://doi.org/10.1016/j.molmed.2020.01.001 [PubMed]
- 148. González-Gualda E, Pàez-Ribes M, Lozano-Torres B, Macias D, Wilson JR
3rd , González-López C, Ou HL, Mirón-Barroso S, Zhang Z, Lérida-Viso A, Blandez JF, Bernardos A, Sancenón F, et al. Galacto-conjugation of Navitoclax as an efficient strategy to increase senolytic specificity and reduce platelet toxicity. Aging Cell. 2020; 19:e13142. https://doi.org/10.1111/acel.13142 [PubMed] - 149. Kang C. Senolytics and Senostatics: A Two-Pronged Approach to Target Cellular Senescence for Delaying Aging and Age-Related Diseases. Mol Cells. 2019; 42:821–27. https://doi.org/10.14348/molcells.2019.0298 [PubMed]
- 150. Willard-Mack CL. Normal structure, function, and histology of lymph nodes. Toxicol Pathol. 2006; 34:409–24. https://doi.org/10.1080/01926230600867727 [PubMed]
- 151. Bousso P. T-cell activation by dendritic cells in the lymph node: lessons from the movies. Nat Rev Immunol. 2008; 8:675–84. https://doi.org/10.1038/nri2379 [PubMed]
- 152. Capece T, Kim M. The Role of Lymphatic Niches in T Cell Differentiation. Mol Cells. 2016; 39:515–23. https://doi.org/10.14348/molcells.2016.0089 [PubMed]
- 153. Woodland DL, Kohlmeier JE. Migration, maintenance and recall of memory T cells in peripheral tissues. Nat Rev Immunol. 2009; 9:153–61. https://doi.org/10.1038/nri2496 [PubMed]
- 154. Liao S, von der Weid PY. Lymphatic system: an active pathway for immune protection. Semin Cell Dev Biol. 2015; 38:83–89. https://doi.org/10.1016/j.semcdb.2014.11.012 [PubMed]
- 155. Ahmadi O, McCall JL, Stringer MD. Does senescence affect lymph node number and morphology? A systematic review. ANZ J Surg. 2013; 83:612–18. https://doi.org/10.1111/ans.12067 [PubMed]
- 156. Hadamitzky C, Spohr H, Debertin AS, Guddat S, Tsokos M, Pabst R. Age-dependent histoarchitectural changes in human lymph nodes: an underestimated process with clinical relevance? J Anat. 2010; 216:556–62. https://doi.org/10.1111/j.1469-7580.2010.01213.x [PubMed]
- 157. Richner JM, Gmyrek GB, Govero J, Tu Y, van der Windt GJ, Metcalf TU, Haddad EK, Textor J, Miller MJ, Diamond MS. Age-Dependent Cell Trafficking Defects in Draining Lymph Nodes Impair Adaptive Immunity and Control of West Nile Virus Infection. PLoS Pathog. 2015; 11:e1005027. https://doi.org/10.1371/journal.ppat.1005027 [PubMed]
- 158. Thompson HL, Smithey MJ, Surh CD, Nikolich-Žugich J. Functional and Homeostatic Impact of Age-Related Changes in Lymph Node Stroma. Front Immunol. 2017; 8:706. https://doi.org/10.3389/fimmu.2017.00706 [PubMed]
- 159. Zhao J, Zhao J, Legge K, Perlman S. Age-related increases in PGD(2) expression impair respiratory DC migration, resulting in diminished T cell responses upon respiratory virus infection in mice. J Clin Invest. 2011; 121:4921–30. https://doi.org/10.1172/JCI59777 [PubMed]
- 160. Basisty N, Kale A, Jeon OH, Kuehnemann C, Payne T, Rao C, Holtz A, Shah S, Sharma V, Ferrucci L, Campisi J, Schilling B. A proteomic atlas of senescence-associated secretomes for aging biomarker development. PLoS Biol. 2020; 18:e3000599. https://doi.org/10.1371/journal.pbio.3000599 [PubMed]
- 161. Masters AR, Hall A, Bartley JM, Keilich SR, Lorenzo EC, Jellison ER, Puddington L, Haynes L. Assessment of Lymph Node Stromal Cells as an Underlying Factor in Age-Related Immune Impairment. J Gerontol A Biol Sci Med Sci. 2019; 74:1734–43. https://doi.org/10.1093/gerona/glz029 [PubMed]
- 162. Denton AE, Silva-Cayetano A, Dooley J, Hill DL, Carr EJ, Robert PA, Meyer-Hermann M, Liston A, Linterman MA. Intrinsic defects in lymph node stromal cells underpin poor germinal center responses during aging. bioRxiv. 2020. https://doi.org/10.1101/2020.05.07.082255
- 163. Dasoveanu DC, Park HJ, Ly CL, Shipman WD, Chyou S, Kumar V, Tarlinton D, Ludewig B, Mehrara BJ, Lu TT. Lymph node stromal CCL2 limits antibody responses. Sci Immunol. 2020; 5:eaaw0693. https://doi.org/10.1126/sciimmunol.aaw0693 [PubMed]
- 164. Randall TD. Bronchus-associated lymphoid tissue (BALT) structure and function. Adv Immunol. 2010; 107:187–241. https://doi.org/10.1016/B978-0-12-381300-8.00007-1 [PubMed]
- 165. Hwang JY, Randall TD, Silva-Sanchez A. Inducible Bronchus-Associated Lymphoid Tissue: Taming Inflammation in the Lung. Front Immunol. 2016; 7:258. https://doi.org/10.3389/fimmu.2016.00258 [PubMed]
- 166. Knop N, Knop E. Conjunctiva-associated lymphoid tissue in the human eye. Invest Ophthalmol Vis Sci. 2000; 41:1270–79. [PubMed]
- 167. Mastropasqua R, Agnifili L, Fasanella V, Nubile M, Gnama AA, Falconio G, Perri P, Di Staso S, Mariotti C. The Conjunctiva-Associated Lymphoid Tissue in Chronic Ocular Surface Diseases. Microsc Microanal. 2017; 23:697–707. https://doi.org/10.1017/S1431927617000538 [PubMed]
- 168. Kracke A, Hiller AS, Tschernig T, Kasper M, Kleemann WJ, Tröger HD, Pabst R. Larynx-associated lymphoid tissue (LALT) in young children. Anat Rec. 1997; 248:413–20. https://doi.org/10.1002/(SICI)1097-0185(199707)248:3<413::AID-AR14>3.0.CO;2-S [PubMed]
- 169. Ono S, Kabashima K. Novel insights into the role of immune cells in skin and inducible skin-associated lymphoid tissue (iSALT). Allergo J Int. 2015; 24:170–79. https://doi.org/10.1007/s40629-015-0065-1 [PubMed]
- 170. Ono S, Kabashima K. Proposal of inducible skin-associated lymphoid tissue (iSALT). Exp Dermatol. 2015; 24:630–31. https://doi.org/10.1111/exd.12716 [PubMed]
- 171. Kuper CF, Koornstra PJ, Hameleers DM, Biewenga J, Spit BJ, Duijvestijn AM, van Breda Vriesman PJ, Sminia T. The role of nasopharyngeal lymphoid tissue. Immunol Today. 1992; 13:219–24. https://doi.org/10.1016/0167-5699(92)90158-4 [PubMed]
- 172. Debertin AS, Tschernig T, Tönjes H, Kleemann WJ, Tröger HD, Pabst R. Nasal-associated lymphoid tissue (NALT): frequency and localization in young children. Clin Exp Immunol. 2003; 134:503–07. https://doi.org/10.1111/j.1365-2249.2003.02311.x [PubMed]
- 173. Jung C, Hugot JP, Barreau F. Peyer’s Patches: The Immune Sensors of the Intestine. Int J Inflam. 2010; 2010:823710. https://doi.org/10.4061/2010/823710 [PubMed]
- 174. Hellings P, Jorissen M, Ceuppens JL. The Waldeyer’s ring. Acta Otorhinolaryngol Belg. 2000; 54:237–41. [PubMed]
- 175. Brandtzaeg P. Immunology of tonsils and adenoids: everything the ENT surgeon needs to know. Int J Pediatr Otorhinolaryngol. 2003 (Suppl 1); 67:S69–76. https://doi.org/10.1016/j.ijporl.2003.08.018 [PubMed]
- 176. Fujimura Y, Akisada T, Harada T, Haruma K. Uptake of microparticles into the epithelium of human nasopharyngeal lymphoid tissue. Med Mol Morphol. 2006; 39:181–86. https://doi.org/10.1007/s00795-006-0335-6 [PubMed]
- 177. Forchielli ML, Walker WA. The role of gut-associated lymphoid tissues and mucosal defence. Br J Nutr. 2005 (Suppl 1); 93:S41–48. https://doi.org/10.1079/bjn20041356 [PubMed]
- 178. McGhee JR, Fujihashi K. Inside the mucosal immune system. PLoS Biol. 2012; 10:e1001397. https://doi.org/10.1371/journal.pbio.1001397 [PubMed]
- 179. Faria AM, Garcia G, Rios MJ, Michalaros CL, Vaz NM. Decrease in susceptibility to oral tolerance induction and occurrence of oral immunization to ovalbumin in 20-38-week-old mice. The effect of interval between oral exposures and rate of antigen intake in the oral immunization. Immunology. 1993; 78:147–51. [PubMed]
- 180. Hagiwara Y, McGhee JR, Fujihashi K, Kobayashi R, Yoshino N, Kataoka K, Etani Y, Kweon MN, Tamura S, Kurata T, Takeda Y, Kiyono H, Fujihashi K. Protective mucosal immunity in aging is associated with functional CD4+ T cells in nasopharyngeal-associated lymphoreticular tissue. J Immunol. 2003; 170:1754–62. https://doi.org/10.4049/jimmunol.170.4.1754 [PubMed]
- 181. Koga T, McGhee JR, Kato H, Kato R, Kiyono H, Fujihashi K. Evidence for early aging in the mucosal immune system. J Immunol. 2000; 165:5352–59. https://doi.org/10.4049/jimmunol.165.9.5352 [PubMed]
- 182. Klingenberg B, Hafkamp HC, Haesevoets A, Manni JJ, Slootweg PJ, Weissenborn SJ, Klussmann JP, Speel EJ. p16 INK4A overexpression is frequently detected in tumour-free tonsil tissue without association with HPV. Histopathology. 2010; 56:957–67. https://doi.org/10.1111/j.1365-2559.2010.03576.x [PubMed]
- 183. Chen S, Wang WW, Wang Y, Li YQ, Zhu LX. Cellular senescence in recurrent tonsillitis and tonsillar hypertrophy in children. Int J Pediatr Otorhinolaryngol. 2020; 133:110004. https://doi.org/10.1016/j.ijporl.2020.110004 [PubMed]
- 184. Yu Y, Park YS, Kim HS, Kim HY, Jin YM, Jung SC, Ryu KH, Jo I. Characterization of long-term in vitro culture-related alterations of human tonsil-derived mesenchymal stem cells: role for CCN1 in replicative senescence-associated increase in osteogenic differentiation. J Anat. 2014; 225:510–18. https://doi.org/10.1111/joa.12229 [PubMed]
- 185. Choi DH, Oh SY, Choi JK, Lee KE, Lee JY, Park YJ, Jo I, Park YS. A transcriptomic analysis of serial-cultured, tonsil-derived mesenchymal stem cells reveals decreased integrin α3 protein as a potential biomarker of senescent cells. Stem Cell Res Ther. 2020; 11:359. https://doi.org/10.1186/s13287-020-01860-y [PubMed]
- 186. Fujihashi K, Kiyono H. Mucosal immunosenescence: new developments and vaccines to control infectious diseases. Trends Immunol. 2009; 30:334–43. https://doi.org/10.1016/j.it.2009.04.004 [PubMed]
- 187. Kawanishi H. Recent progress in senescence-associated gut mucosal immunity. Dig Dis. 1993; 11:157–72. https://doi.org/10.1159/000171408 [PubMed]
- 188. Koyama K, Hosokawa T, Aoike A. Aging effect on the immune functions of murine gut-associated lymphoid tissues. Dev Comp Immunol. 1990; 14:465–73. https://doi.org/10.1016/0145-305x(90)90039-h [PubMed]
- 189. Moretto MM, Lawlor EM, Khan IA. Aging mice exhibit a functional defect in mucosal dendritic cell response against an intracellular pathogen. J Immunol. 2008; 181:7977–84. https://doi.org/10.4049/jimmunol.181.11.7977 [PubMed]
- 190. Kato H, Fujihashi K, Kato R, Dohi T, Fujihashi K, Hagiwara Y, Kataoka K, Kobayashi R, McGhee JR. Lack of oral tolerance in aging is due to sequential loss of Peyer’s patch cell interactions. Int Immunol. 2003; 15:145–58. https://doi.org/10.1093/intimm/dxg011 [PubMed]
- 191. Sreekumar PG, Hinton DR, Kannan R. The Emerging Role of Senescence in Ocular Disease. Oxid Med Cell Longev. 2020; 2020:2583601. https://doi.org/10.1155/2020/2583601 [PubMed]
- 192. Bartling B. Cellular senescence in normal and premature lung aging. Z Gerontol Geriatr. 2013; 46:613–22. https://doi.org/10.1007/s00391-013-0543-3 [PubMed]
- 193. Campisi J. The role of cellular senescence in skin aging. J Investig Dermatol Symp Proc. 1998; 3:1–5. [PubMed]
- 194. Penfield JD, Anderson M, Lutzke L, Wang KK. The role of cellular senescence in the gastrointestinal mucosa. Gut Liver. 2013; 7:270–77. https://doi.org/10.5009/gnl.2013.7.3.270 [PubMed]
- 195. Liu Y, Bloom SI, Donato AJ. The role of senescence, telomere dysfunction and shelterin in vascular aging. Microcirculation. 2019; 26:e12487. https://doi.org/10.1111/micc.12487 [PubMed]
- 196. Minamino T, Komuro I. Vascular cell senescence: contribution to atherosclerosis. Circ Res. 2007; 100:15–26. https://doi.org/10.1161/01.RES.0000256837.40544.4a [PubMed]
- 197. Fukuiwa T, Sekine S, Kobayashi R, Suzuki H, Kataoka K, Gilbert RS, Kurono Y, Boyaka PN, Krieg AM, McGhee JR, Fujihashi K. A combination of Flt3 ligand cDNA and CpG ODN as nasal adjuvant elicits NALT dendritic cells for prolonged mucosal immunity. Vaccine. 2008; 26:4849–59. https://doi.org/10.1016/j.vaccine.2008.06.091 [PubMed]
- 198. Dong Y, Lian X, Xu Y, Hu H, Chang C, Zhang H, Zhang L. Hematopoietic stem/progenitor cell senescence is associated with altered expression profiles of cellular memory-involved gene. Biosci Rep. 2018; 38:BSR20171589. https://doi.org/10.1042/BSR20171589 [PubMed]
- 199. Kim HN, Chang J, Shao L, Han L, Iyer S, Manolagas SC, O'Brien CA, Jilka RL, Zhou D, Almeida M. DNA damage and senescence in osteoprogenitors expressing Osx1 may cause their decrease with age. Aging Cell. 2017; 16:693–703. https://doi.org/10.1111/acel.12597 [PubMed]
- 200. Nakamura-Ishizu A, Suda T. Aging of the hematopoietic stem cells niche. Int J Hematol. 2014; 100:317–25. https://doi.org/10.1007/s12185-014-1641-8 [PubMed]
- 201. Palmer DB. The effect of age on thymic function. Front Immunol. 2013; 4:316. https://doi.org/10.3389/fimmu.2013.00316 [PubMed]
- 202. Turner VM, Mabbott NA. Structural and functional changes to lymph nodes in ageing mice. Immunology. 2017; 151:239–47. https://doi.org/10.1111/imm.12727 [PubMed]