



Introduction
Young children who experienced multiple surgical procedures under general anesthesia before 3 years of age are likely to fall within a high-risk category as defined by the recent US Food and Drug Administration warning: “repeated anesthesia in surgery or lengthy use of sedatives and general anesthetics (> 3 hours) may affect the brain development in children < 3 years” [1–4]. However, single and short-duration anesthesia and surgery do not cause a detectable impact on neurodevelopment in children [5, 6]. Remarkably, a recent prospective study revealed that children receiving multiple exposures to anesthesia show problems in behaviors, executive function, and reading [7]. Thus, exposure to anesthetics during childhood may lead to long-term deficits in neurodevelopmental function of children [8–10].
Sevoflurane, which has the favorable properties of low pungency, rapid onset, rapid offset, and low blood/gas ratio, is the most commonly used inhalational anesthetic agent for pediatric patients [11, 12]. Numerous preclinical studies have shown that repeated sevoflurane exposure during the neonatal period causes cognitive impairment and neurotoxicity in young rodents and monkeys [13–17], which may be associated with nerve cell apoptosis [18], impairment of synaptic development [19], neurogenesis inhibition [20], or impairment of glial cells development [21]. In addition, treatments and possible mechanisms for cognitive impairment caused by repeated sevoflurane exposure have been extensively studied in the developing central nervous system. However, the underlying mechanisms of peripheral responses on cognitive impairment in young mice after multiple neonatal exposures to sevoflurane remain elusive.
The gut microbiota is a peripheral microorganism community in the digestive tract that helps to maintain dynamic metabolic ecological balance [22, 23]. The gut microbiota, not only through the nervous system (the gut–brain neuroanatomical pathway) but also through the endocrine system, immune system, and metabolic system, affords significant advantages to the host throughout development [24]. However, abnormal composition of the gut microbiota can substantially influence the function and microenvironment of the brain [25–28], through the three routes of the gut–brain axis (immune, neuroendocrine, and vagal nerve pathway) [28–30]. Mounting evidence indicates that inhalation anesthesia can alter the composition of the gut microbiota in mice [31]. Recent studies have revealed significant interactions between alteration of the gut microbiota and cognitive behavior [32, 33]. However, scant attention has been paid to investigating the relationship between cognitive impairment and abnormal composition of the gut microbiota in young mice after multiple neonatal exposures to sevoflurane.
Therefore, we used a mouse model of multiple neonatal exposures to sevoflurane to evaluate the effects of anesthesia on cognitive function. Sequencing of the 16S ribosomal RNA gene and analysis of fecal samples were used to assess changes in the gut microbiota. In addition, we examined the effects of fecal microbiota transplantation on cognitive behavior in antibiotic-induced pseudo germ-free mice. The primary objective was to determine whether multiple neonatal exposures to sevoflurane impact on the gut microbiota of the young mice to induce cognitive impairment. The findings will contribute to an improved understanding of the mechanisms of anesthesia-related neurotoxicity.
Results
Comparison of cognitive behavior between anesthesia group and control group
The study design is summarized in Figure 1A. The mice from the anesthesia group (i.e., receiving 3% sevoflurane three times on PND (postnatal day) 6, 8, and 10 (for 2 h at each application) exhibited longer escape latency in the MWM (Morris water maze) test during the training phase on PND 33–35 (Figure 1D). In addition, the anesthesia group exhibited reduced spatial positioning capability as demonstrated by the results from the probe trial on PND 36, including showing fewer platform-crossing instances (4.80 ± 0.53 vs 2.20 ± 0.49, p = 0.0021; Figure 1E), less time spent in the fourth quadrant (26.10 ± 1.70 s vs 18.31 ± 2.06 s, p = 0.0092; Figure 1F), and longer mean distance from the platform (0.27 ± 0.02 m vs 0.35 ± 0.02 m, p = 0.0041; Figure 1G) compared with those of the control group. These results suggested that the sevoflurane treatment may result in defective memory and cognitive function of the treated mice [19, 34].
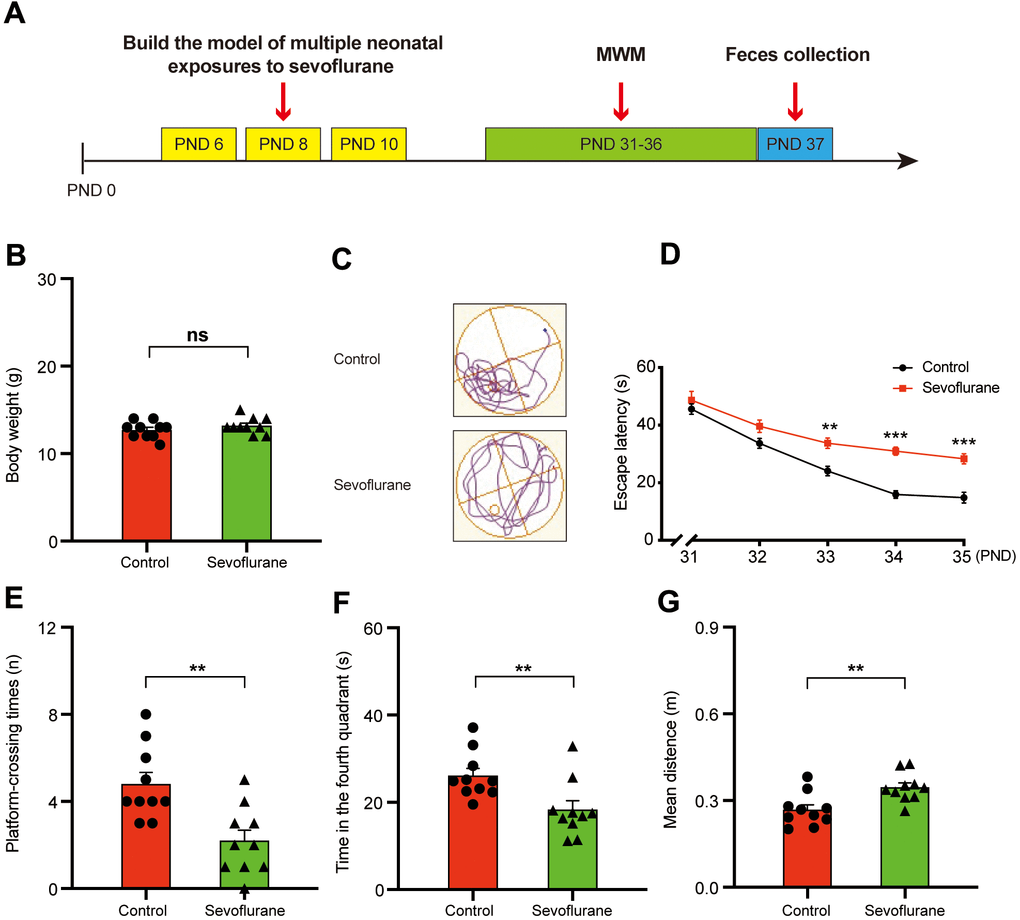
Figure 1. Morris water maze test for control and sevoflurane-treated mice. (A) Experimental schedule: 3% sevoflurane was applied for 2 h daily on PND 6, 8, and 10, MWM test on PND 31–36, and fecal sample collection for 16S ribosomal RNA gene sequencing and fecal bacteria transplant on PND 37. (B) Body weight (Student’s t-test, p > 0.05). (C) Trace plot of control and sevoflurane-treated mice in the MWM test. (D) Escape latency (two-way ANOVA; Time: F4,72 = 67.43, p < 0.001; Group: F1,18 = 43.14, p < 0.001; Interaction: F4,72 = 3.857, p = 0.007). (E) Platform-crossing instances (Student’s t-test, p = 0.0021). (F) Time spent in the fourth quadrant (Student’s t-test, p = 0.0092). (G) Mean distance from the platform (Student’s t-test, p = 0.0041). PND: postnatal day; ANOVA: analysis of variance; MWM: Morris water maze. Data are shown as mean ± SEM (n = 10). Significance: * p < 0.05, ** p < 0.01, *** p < 0.001, ns: non-significant.
Alterations in the gut microbiota composition between control group and anesthesia group
Recent studies have demonstrated that certain microorganisms in the gut microbiota play critical roles in the memory and cognitive behaviors of the hosts [30, 35]. We sought to determine whether the cognitive behavioral difference between the control and sevoflurane-treated mice was associated with a change in the gut microbiota. Given that body weight has a strong impact on the gut microbiota [36], we first measured the body weight of the control and sevoflurane-treated mice. No difference in body weight was observed between the two groups (12.70 ± 0.30 g vs 13.20 ± 0.29 g, p = 0.25; Figure 1B).
Fecal samples were then collected for 16S ribosomal RNA gene sequencing. Based on the comprehensive PKSSU4.0 database implemented in the EzBioCloud platform, the bacterial taxonomic composition of the control and sevoflurane-treated mice was determined and compared. No significant difference in alpha-diversity between the two groups was observed, as indicated by the Shannon and Simpson indices (p > 0.05). Principal coordinate analysis indicated that the bacterial communities of the anesthesia group were not significantly different from those of the control group. These results suggested that sevoflurane treatment did not significantly alter the overall taxonomic composition of the gut microbiota. Previous reports also suggested that cognitive dysfunction caused by streptozotocin-induced diabetes or high-cholesterol diet did not significantly alter the gut microbiota composition in mice [37, 38]. However, we observed that multiple taxa exhibited significant differences in relative abundance between the sevoflurane-treated and control mice (Supplementary Table 1 and Figure 2). Among these taxa, several have been reported to be associated with memory and/or health of the hosts. For instance, the genus Streptococcus, which is an important pathogen causing neural damage and a risk factor for cerebral microbleeds and cognitive impairment [39, 40], exhibited increased relative abundance in the sevoflurane group. In contrast, the species PAC001381_s, an uncultured taxon belonging to the Lachnospiraceae, exhibited decreased relative abundance in sevoflurane-treated mice. Lachnospiraceae is an important member of the gut microbiota that can benefit the host in multiple aspects by producing beneficial metabolites, such as propionate and tryptophan [41]. Decreased Lachnospiraceae abundance in the gut microbiota were associated with a worse clinical profile, including higher frequencies of cognitive impairment [42]; while higher Lachnospiraceae abundance were associated with good cognition behaviors independent of clinical variables [43]. Decreased relative abundance of two species affiliated with Pseudoflavonifractor, PAC002302_s and PAC002490_s, in the sevoflurane-treated mice was observed. Pseudoflavonifractor is positively associated with weight loss of obese patients and may benefit the host by producing short-chain fatty acids [44, 45]. The results suggested that the sevoflurane treatment showed a long-term impact on the gut microbiota, at least 27 days post the sevoflurane treatment as demonstrated by the microbiota profiling results.
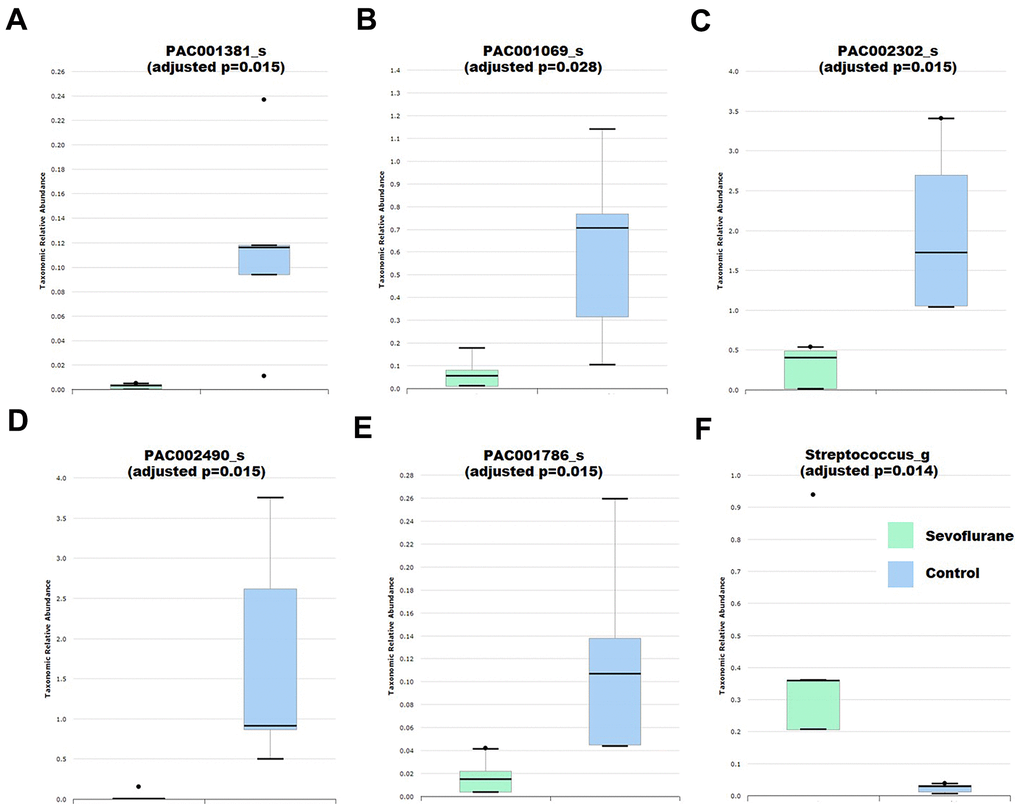
Figure 2. Differential abundance of gut bacteria between control (n = 5) and anesthesia (n = 4) mice. (A) Species PAC001381_s. (B) Species PAC001069_s. (C) Species PAC002302_s. (D) Species PAC002490_s. (E) Species PAC001786_s. (F) Genus Streptococcus. Taxa were assigned based on the PKSSU4.0 database implemented in the EzBioCloud platform.
Effects of control group and anesthesia group gut microbiota transplant on MWM behavior in antibiotic-induced pseudo germ-free mice
To examine whether the defective memory and cognitive function of the sevoflurane-treated mice were due to alteration of the gut microbiota composition, we performed microbiota transplantation experiments using the microbiota from the sevoflurane-treated mice and the control mice as donors and pseudo germ-free mice as recipients. The study design is summarized in Figure 3A. The pseudo germ-free mouse model was established by administering antibiotics at high doses for 14 consecutive days on PND 21–34. Gut microbiota from the control group and anesthesia group were transplanted into the gastrointestinal tract of pseudo germ-free mice for an additional 14 consecutive days on PND 35–48.
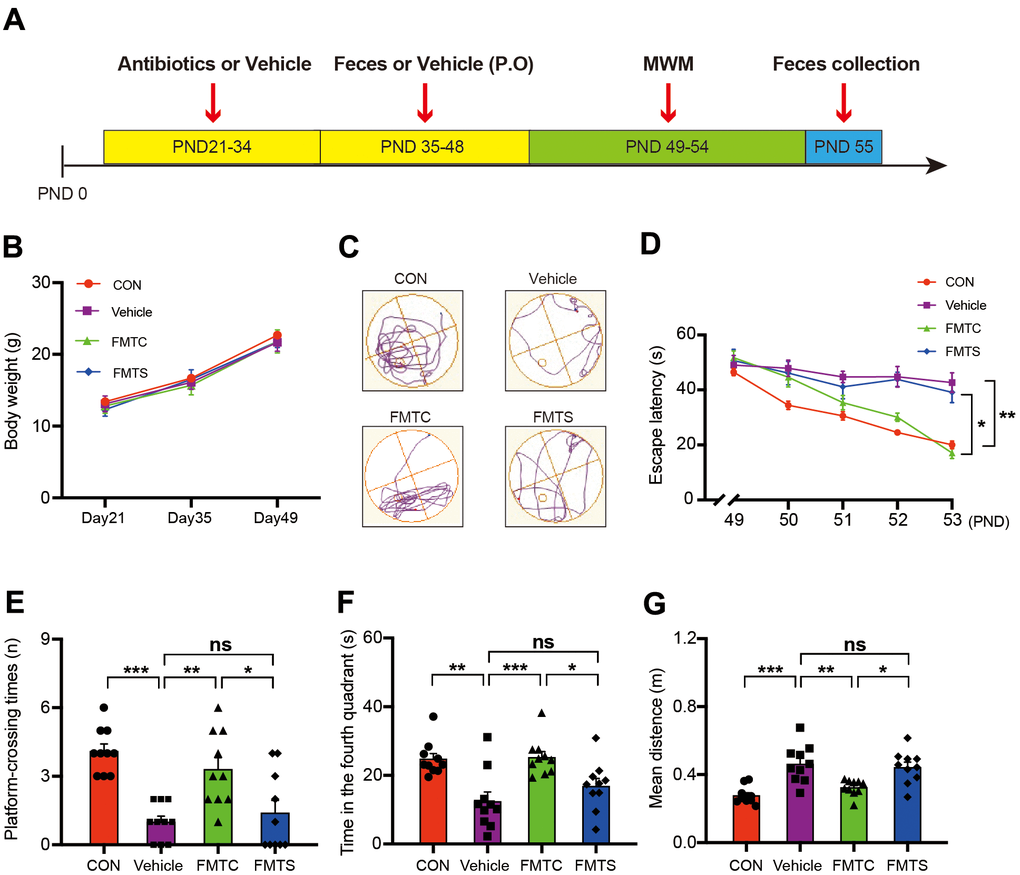
Figure 3. Effects of transplantation of fecal microbiota from control and sevoflurane-treated mice on behavior of pseudo germ-free mice. (A) Experimental summary: fecal microbiota transplantation effects on behavioral testing in pseudo germ-free mice. Wild-type male mice were first treated by administering high doses of antibiotic solution for 14 consecutive days on PND 21–34. Thereafter, mice were orally treated with fetal microbiota of control and anesthesia mice on PND 35–48. The MWM test was performed on PND 49–54. Fecal samples were collected for 16S ribosomal RNA gene sequencing testing on PND 55. (B) Body weight (two-way ANOVA; Time: F2,72 = 959.6, p < 0.001; Group: F3,36 = 1.795, p = 0.17; Interaction: F6,72 = 1.209, p = 0.31). (C) Trace plot of mice in the MWM test. (D) Escape latency (two-way ANOVA; Time: F4,144 = 35.46, p < 0.001; Group: F3,36 = 14.51, p < 0.001; Interaction: F12,144 = 4.436, p < 0.001). (E) Platform-crossing instances (one-way ANOVA; F3,36 = 12.20, p < 0.001). (F) Time spent in the fourth quadrant (one-way ANOVA; F3,36 = 8.812, p = 0.0002). (G) Mean distance from the platform (one-way ANOVA; F3,36 = 12.56, p < 0.001). PND: postnatal day; ANOVA: analysis of variance; MWM: Morris water maze. Data are shown as mean ± SEM (n = 10). * p < 0.05, ** p < 0.01, *** p < 0.001.
No obvious differences in body weight gain among the control (no treatment), pseudo germ-free mice (vehicle), pseudo germ-free mice with transplanted microbiota from the control mice used in the aforementioned experiment (FMTC), and pseudo germ-free mice with transplanted microbiota from the sevoflurane-treated mice used in the aforementioned experiment (FMTS) were observed (Figure 3B). Interestingly, the vehicle mice exhibited increased escape latency time during the training phase of the MWM test (16.31 ± 3.83 s vs 42.70 ± 11.25 s, p = 0.005 for PND 54; Figure 3D), and fewer platform-crossing instances (4.10 ± 0.31 vs 1.00 ± 0.26, p < 0.001; Figure 3E), less time spent in the fourth quadrant (24.80 ± 1.59 s vs 12.45 ± 2.71 s, p = 0.0011; Figure 3F), and longer mean distance from the platform (0.28 ± 0.02 m vs 0.46 ± 0.03 m, p < 0.001; Figure 3G) compared with those of the control group. The defective memory and cognitive function of the vehicle mice were recovered by regaining the regular full-spectrum microbiota (i.e., that of the FMTC mice); however, the microbiota derived from sevoflurane-treated mice (i.e., the FMTS mice) did not dramatically improve the cognitive function of the recipients as demonstrated by the MWM test (Figure 3D–3G).
Effects of fecal microbiota transplant from anesthesia and control mice on the abundance of host gut microbiota
Since all mice used for FMTS and FMTC treatments were on the same mental and gut microbiota composition background (pseudo germ-free vehicle mice), thus the cognitive behavior differences between the FMTS and FMTC mice were not due to the mental status or the original gut microbiota of the used mice, but were mainly due to the transplanted gut microbiota. Therefore, we performed the gut microbiota comparison analysis. The results demonstrated that the microbiota composition of the vehicle mice was very simple with majority of the members in the normal gut microbiota eliminated, while the microbiota composition of the FMTS and FMTC mice was dramatically different from that of the vehicle mice, suggesting the FMT experiments were successfully performed (Supplementary Figure 1). Following an identical bioinformatics analysis protocol, the microbiota composition of FMTS and FMTC mice was compared and the taxa differing significantly in abundance were identified (Supplementary Table 2 and Figure 4). A fraction of differentially abundant taxa identified in the FMTS vs FMTC experiment not overlapping with the abovementioned control vs sevoflurane experiment, which is frequently observed in fecal microbiota transplantation experiments [46]. However, it was interesting to note that several taxa showed consistency between the pre- and post-transplantation microbiota. Control or sevoflurane-treated fecal microbiota transplantation significantly improved or further aggravated the same changes in pseudo germ-free mice (Figure 4A–4F).
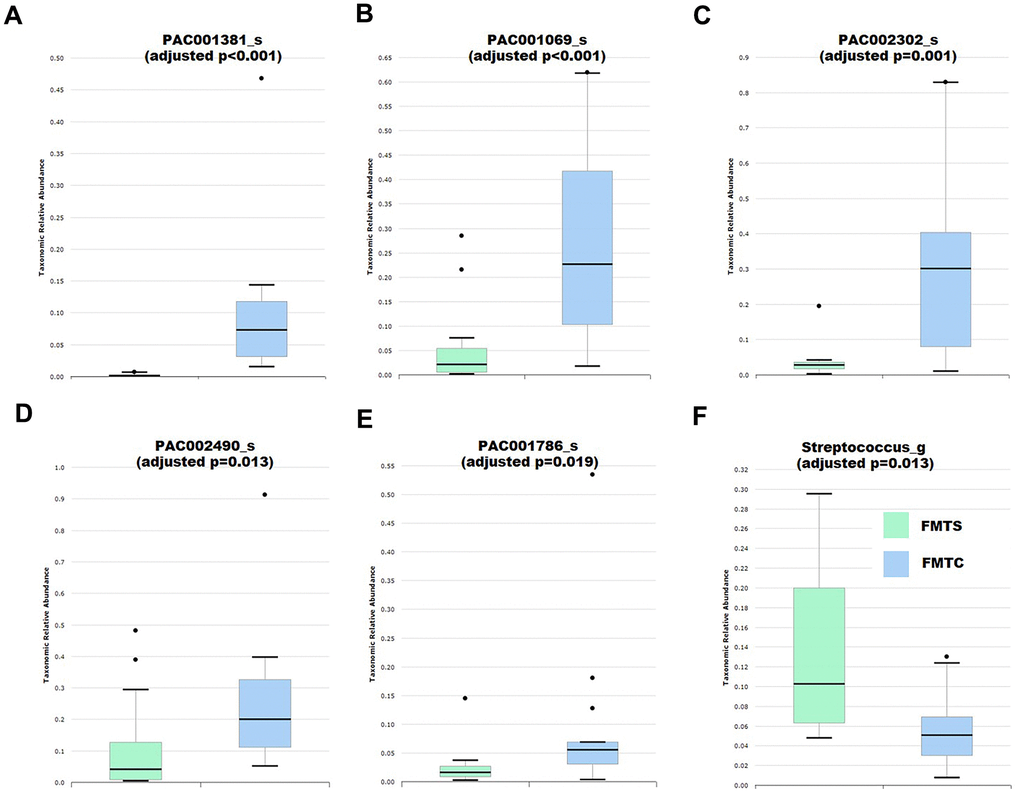
Figure 4. Effects of fecal microbiota transplantation from anesthesia-treated (FMTS, n = 14) and control (FMTC, n = 13) mice on the composition of the host gut microbiota of pseudo-germ-free mice. (A) Species PAC001381_s. (B) Species PAC001069_s. (C) Species PAC002302_s. (D) Species PAC002490_s. (E) Species PAC001786_s. (F) Genus Streptococcus. Taxa were assigned based on the PKSSU4.0 database implemented in the EzBioCloud platform.
Discussion
The newborn period is an extremely important stage for brain development [47, 48]. For rodents, this critical neurodevelopmental process, also termed “the window of vulnerability”, is likely to occur during PND 7–30 when the neuronal architecture and brain function are dramatically trained and improved by environmental events, which may better equip an individual to cope with environmental challenges [49, 50]. Negative environmental factors experienced by an individual during this window of vulnerability can also lead to adverse reactions in brain development [51]. Consistent with the findings of previous studies [52, 53], the present results demonstrated that multiple neonatal exposures to sevoflurane induced cognitive impairment of the young host. However, the mechanisms underlying this anesthesia-triggered cognitive impairment remain elusive.
The present results demonstrated that those sevoflurane-treated neonatal mice that showed learning disability and memory impairment exhibited an altered gut microbiota composition compared with that of the control mice (Figure 2 and Supplementary Table 1). The fecal transplantation experiments using pseudo germ-free mice further demonstrated that the gut microbiota alternation is responsible for the learning disability and memory impairment of the host [54–56]. Comparison of microbiota profiles between the control and sevoflurane-treated mice (first experimental cycle) as well as the FMTC and FMTS mice (second experimental cycle) identified multiple taxa that exhibited consistent differential relative abundance (Figures 2, 4, Supplementary Tables 1, 2). Several of these taxa, such as Lachnospiraceae and Streptococcus, have been reported to be associated with learning and memory functions of the hosts [39, 40, 42, 43]. Accumulating evidence indicates that inhalational anesthetics can affect the process of microbial colonization [31, 57]. The current results further suggested that the memory and cognitive function impairment of the sevoflurane-treated host is due to, at least partially, altered relative abundance of certain crucial members of the gut microbiota.
Recent studies have demonstrated that the gut microbiota is an environmental factor that strongly impacts on brain development and behavior [23, 58, 59]. The gut microbiota is a relatively stable ecosystem; however, the composition and diversity of the gut microbiota is always in a dynamic state during the neonatal period [23]. Newborn babies’ intestines are rapidly colonized by an array of microbes from their mothers, which is characterized by low diversity; in addition, infants possess a stable gut microbial profile that is highly similar to the characteristic microbiota of an adult by the end of the third year of life [60–62]. Therefore, the first 3 years of life represent the most critical period for establishment of the gut microbiota to improve child growth and neurodevelopment, during which time the composition and diversity of the gut microbiota is readily affected by harmful environmental factors, such as bacterial infections and antibiotic treatment [63]. In this study, we found the pseudo germ-free mice exhibited cognitive impairment symptoms. Although it was hard to differentiate and quantify the roles of high dose antibiotics and the consequent gut microbiota alteration on the observed cognitive impairment in the pseudo germ-free mice [64], however, fecal microbiota transplanted from control mice, but not from sevoflurane-treated mice, reversed the detrimental effects on cognitive function in the pseudo germ-free mice, further demonstrating that the microbiota alternation triggered by sevoflurane treatment was responsible for the cognitive impairment in the sevoflurane-treated mice. Taken together, these findings show that multiple neonatal exposures to sevoflurane may strongly impact on microbial colonization of the gut, resulting in altered cognitive phenotypes in adulthood.
Several aspects were not addressed by the present study. First, we primarily focused on the microbial taxa that showed a consistent trend before and after fecal microbiota transplantation to investigate the mechanism of cognitive impairment, but other members of the gut microbiota might also be associated with cognitive function. Second, the microbial alpha-diversity failed to show an apparent difference in response to sevoflurane treatment. Although alpha-diversity is used as a measure of the diversity and richness of the unique microbial taxa within a sample, it should be emphasized that alpha-diversity may not be the sole criterion for gut microbiota dysbiosis [65]. Furthermore, some instances of gut microbiota dysbiosis show no changes in alpha-diversity [66]. Third, the germ-free mouse construct provides an optimal means of assessing long-term effects of specific bacteria on cognitive impairment, but the behavioral assessment of these mice is restricted to sterile isolator units to maintain the germ-free environment. Thus, pseudo germ-free mice provide a more amenable and cost-effective model for behavioral assessment. Finally, under the present experimental design, we observed elements of the gut microbiota that may participate in regulation of cognitive functions in young mice after multiple neonatal exposures to sevoflurane, further investigation is needed to elucidate the roles and the mechanisms of these bacteria on the cognitive functions, and how the microbiota alternation affects the brain tissue to cause the cognitive impairment in the sevoflurane-treated mice.
To the best of our knowledge, the present study is the first to show the effects of an altered gut microbiota on cognitive impairment in young mice after multiple neonatal exposures to sevoflurane. These findings indicate that neonatal exposures to sevoflurane may lead to abnormal composition of the gut microbiota, which is a potential risk factor for neurological development. With its novel insight into sevoflurane-related neurotoxicity in the developing brain, this study presents a promising foundation for further research on microbial manipulation to improve the safety of anesthesia care in children.
Materials and Methods
Animals and anesthesia
The animal studies (ethical protocol number: 201910A204) were conducted in accordance with the guidelines and regulations of the Institutional Animal Care and Use Committee of Soochow University (Suzhou, China). C57BL/6J mice were purchased from the Shanghai Laboratory Animal Center (Shanghai, China) and housed in a specific pathogen-free animal room supplied with standard rodent food and water. The male pups were used in this study. The neonatal mice were randomly assigned to either of the two study groups (control or sevoflurane treatment) with the aid of a computer-generated table.
The animal model was described in our previous study [52]. The mice in the sevoflurane group received 3% sevoflurane with 60% oxygen (balanced with nitrogen) for three non-consecutive days (postnatal days [PND] 6, 8, and 10), for 2 h at each application (2 L/min fresh gas from the start up to 3 min for induction, followed by 1 L/min for maintenance) in a chamber using a Datex-Ohmeda anesthesia system (Madison, WI, USA), which conceptually mimics the multiple exposures of anesthesia in patients [67]. The control group received 60% oxygen in nitrogen for 2 h with an equal rate of flow in a chamber that was identical to the anesthesia chamber [13]. The sevoflurane concentration was continuously monitored using a gas analyzer (Vamos; Dräger Medical, Lübeck, Germany) during the anesthesia. The rectal temperature of the mice was maintained at 37 ± 0.5° C. After anesthesia, the mice were returned to home cages (metabolic cage: allowing feces to leak out to prevent eating each other's feces) under standard care [53, 68]. Given that previous studies demonstrated that anesthesia with 3% sevoflurane did not significantly change the pH, partial pressures of oxygen, partial pressures of carbon dioxide, and hematocrit of the young mice [52], we did not perform blood gas analysis of the mice in this study.
Morris water maze (MWM) Test
A Morris water maze (MWM) test was conducted as previously described [69]. The water maze device, i.e., a round steel pool (150 cm diameter and 60 cm height) with a 10 cm diameter-size platform located in the center, was surrounded by a black curtain and located in an isolated, quiet room. The device was filled with water to a level 1.0 cm above the surface of the platform. Throughout the experiment, the water temperature was maintained at 22° C. The mice were trained to reach the platform for five consecutive days (PND 31–35) with four trials per day. In the training phase each mouse was placed in the water and given 60 s to locate the platform; if the mice could not find the platform within 60 s, they were gently guided to the platform and allowed to remain there for 15 s. The time and routine for each mouse to reach the platform was recorded by video-tracking software (ANY-maze, Stoelting, CO, USA) to evaluate its spatial learning ability. The platform was removed in the testing phase on PND 36, then a 60 s probe trial was performed for assessment of memory function. The number of times the mice crossed the platform area and time spent in the fourth quadrant were recorded. The mice were warmed and dried with a heat lamp after each test.
Pseudo germ-free mice model establishment
Pseudo germ-free mice were established as described by previous studies [51, 70] with slight modification. Briefly, C57BL/6J male mice were treated with a broad-spectrum antibiotics cocktail (ampicillin 1 g/L, neomycin sulfate 1 g/L, and metronidazole 1 g/L; Sigma-Aldrich, St Louis, MO, USA) in drinking water for 14 consecutive days. The drinking solution was renewed every 2 days.
Fecal microbiota transplant and MWM test
The male mice were placed in a clean cage after the MWM test. The 30 male mice were divided into the sevoflurane group (15) and control group (15). Ten mice in each group were prepared for fecal microbiota transplantation and the remaining mice in each group were used for 16S rDNA sequencing. The mice were killed on day 37 and the feces samples were collected from the ileocecal region and placed in a sterilized centrifuge tube. Feces were stored in a −80° C freezer until analysis and transplantation [71]. The fecal microbiota was prepared by diluting 1 g of the fecal sample obtained from the anesthesia group or control group mice in 10 mL sterile PBS. The fecal material was suspended and 0.2 mL of the suspension was guided by gavage into each mouse recipient for 14 consecutive days, and the pseudo germ-free mice were used as the recipients [70]. The MWM test was performed for the control (i.e., untreated), vehicle mice (i.e., pseudo germ-free), FMTC (i.e., pseudo germ-free mice with transplanted microbiota from the control mice used in the aforementioned experiment), and FMTS (i.e., pseudo germ-free mice with transplanted microbiota from the sevoflurane-treated mice used in the aforementioned experiment) as described in section 2.2.
High-throughput 16S rDNA sequencing of the fecal samples and bioinformatics analyses
The fecal samples from the control and sevoflurane-treated mice used in the first cycle experiment, as well as the FMTC, and FMTS and vehicle mice used in the second cycle, were collected immediately from the ileocecal region after the MWM test (Figures 1A, 3A). Samples were placed in 1.5 ml tubes, snap-frozen on dry ice, and stored at −80° C. The 16S rDNA high-throughput sequencing was performed by Sangon Biotech Co., Ltd (Shanghai, China). DNA was extracted using EZNA Soil DNA Kits (Omega, Doraville, GA, USA). The 16S rDNA V3–V5 region was amplified with the primer set 338F (5′-ACTCCTACGGGAGGCAGC-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′). Triplicate PCR reactions for each sample were performed and merged for sequencing. The reverse primer contained a sample barcode and both primers were connected with an Illumina sequencing adapter. The PCR products were purified and sequenced on an Illumina MiSeq PE300 platform. Original sequencing data from the sample were sorted by unique barcodes, followed by removal of the barcode, linker, and PCR primer sequences. The resultant paired-end sequencing data were merged using FLASH [72], and the merged fastq files were analyzed using the 16S rRNA gene-based Microbiome Taxonomic Profiling pipeline implemented on the EzBioCloud server (https://www.ezbiocloud.net/) with the EzBioCloud 16S rDNA database version PKSSU4.0 employed [73]. Comparisons of the taxonomic composition between the control and the sevoflurane-treated mice (named as Sevoflurane herein) or the FMTC and FMTS mice using the comparative MTP analyzer implemented in EzBioCloud, and the alpha- and beta-diversity between the control and Sevoflurane groups or FMTC and FMTS groups, were conducted using normalized data with variation in the gene copy number considered. The taxonomic biomarkers for each group were identified using the linear discriminant analysis effect size (LEfSe) algorithm [74] using the aforementioned normalized data; the taxa with an adjusted p-value (false discovery rate) < 0.05 were considered to be significantly group-associated biomarkers.
The sequencing data have been deposited in the CNGB CNSA database under the Bioproject accession no. CNP0001462.
Statistical analysis
Values presented are expressed as the mean ± SEM. Statistical analyses were performed using SPSS version 17.0 software (SPSS Inc., Chicago, IL, USA). Escape path length and escape latency in the MWM test were analyzed using two-way analysis of variance (ANOVA). Other data were analyzed using one-way ANOVA followed by post-hoc Tukey’s test, Student’s t-test, or Fisher’s exact test. P-values less than 0.05 were considered statistically significant.
Author Contributions
FJ and ML conceived and designed this project. SS, YZ, QC, WC performed the experiments, conducted statistical analyses, and generated the figures. ML and FJ wrote the first draft of the manuscript. YZ, SS and JS revised the manuscript. All the authors approved the final manuscript for submission.
Acknowledgments
We thank Robert McKenzie, PhD, from Liwen Bianji, Edanz Editing China (www.liwenbianji.cn/ac), for editing the English text of a draft of this manuscript.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Funding
This work was supported by the National Natural Science Foundation of China (82072130 and 81873925 to FHJ), Natural Science Foundation of Jiangsu Province (BK20191171 to FHJ), and Science and Technology Development Plan Clinical Trial Project (SLT201909 to FHJ).
Editorial Note
This corresponding author has a verified history of publications using a personal email address for correspondence
References
- 1. Davidson A, Vutskits L. The new FDA drug safety communication on the use of general anesthetics in young children: what should we make of it? Paediatr Anaesth. 2017; 27:336–37. https://doi.org/10.1111/pan.13122 [PubMed]
- 2. Shi Y, Hu D, Rodgers EL, Katusic SK, Gleich SJ, Hanson AC, Schroeder DR, Flick RP, Warner DO. Epidemiology of general anesthesia prior to age 3 in a population-based birth cohort. Paediatr Anaesth. 2018; 28:513–19. https://doi.org/10.1111/pan.13359 [PubMed]
- 3. Vinson AE, Houck CS. Neurotoxicity of Anesthesia in Children: Prevention and Treatment. Curr Treat Options Neurol. 2018; 20:51. https://doi.org/10.1007/s11940-018-0536-z [PubMed]
- 4. Oba S, Işıl CT, Türk HŞ, Karamürsel S, Aksu S, Kaba M, Kılınç L, Dokucu AI. Evaluation of Neurotoxicity of Multiple Anesthesia in Children Using Visual Evoked Potentials. Sisli Etfal Hastan Tip Bul. 2019; 53:284–89. https://doi.org/10.14744/SEMB.2018.59454 [PubMed]
- 5. Sun LS, Li G, Miller TL, Salorio C, Byrne MW, Bellinger DC, Ing C, Park R, Radcliffe J, Hays SR, DiMaggio CJ, Cooper TJ, Rauh V, et al. Association Between a Single General Anesthesia Exposure Before Age 36 Months and Neurocognitive Outcomes in Later Childhood. JAMA. 2016; 315:2312–20. https://doi.org/10.1001/jama.2016.6967 [PubMed]
- 6. Davidson AJ, Disma N, de Graaff JC, Withington DE, Dorris L, Bell G, Stargatt R, Bellinger DC, Schuster T, Arnup SJ, Hardy P, Hunt RW, Takagi MJ, et al, and GAS Consortium. Neurodevelopmental outcome at 2 years of age after general anaesthesia and awake-regional anaesthesia in infancy (GAS): an international multicentre, randomised controlled trial. Lancet. 2016; 387:239–50. https://doi.org/10.1016/S0140-6736(15)00608-X [PubMed]
- 7. Warner DO, Zaccariello MJ, Katusic SK, Schroeder DR, Hanson AC, Schulte PJ, Buenvenida SL, Gleich SJ, Wilder RT, Sprung J, Hu D, Voigt RG, Paule MG, et al. Neuropsychological and Behavioral Outcomes after Exposure of Young Children to Procedures Requiring General Anesthesia: The Mayo Anesthesia Safety in Kids (MASK) Study. Anesthesiology. 2018; 129:89–105. https://doi.org/10.1097/ALN.0000000000002232 [PubMed]
- 8. Backeljauw B, Holland SK, Altaye M, Loepke AW. Cognition and Brain Structure Following Early Childhood Surgery With Anesthesia. Pediatrics. 2015; 136:e1–12. https://doi.org/10.1542/peds.2014-3526 [PubMed]
- 9. Vutskits L, Xie Z. Lasting impact of general anaesthesia on the brain: mechanisms and relevance. Nat Rev Neurosci. 2016; 17:705–17. https://doi.org/10.1038/nrn.2016.128 [PubMed]
- 10. Glatz P, Sandin RH, Pedersen NL, Bonamy AK, Eriksson LI, Granath F. Association of Anesthesia and Surgery During Childhood With Long-term Academic Performance. JAMA Pediatr. 2017; 171:e163470. https://doi.org/10.1001/jamapediatrics.2016.3470 [PubMed]
- 11. Gibert S, Sabourdin N, Louvet N, Moutard ML, Piat V, Guye ML, Rigouzzo A, Constant I. Epileptogenic effect of sevoflurane: determination of the minimal alveolar concentration of sevoflurane associated with major epileptoid signs in children. Anesthesiology. 2012; 117:1253–61. https://doi.org/10.1097/ALN.0b013e318273e272 [PubMed]
- 12. Edwards DA, Shah HP, Cao W, Gravenstein N, Seubert CN, Martynyuk AE. Bumetanide alleviates epileptogenic and neurotoxic effects of sevoflurane in neonatal rat brain. Anesthesiology. 2010; 112:567–75. https://doi.org/10.1097/ALN.0b013e3181cf9138 [PubMed]
- 13. Lu H, Liufu N, Dong Y, Xu G, Zhang Y, Shu L, Soriano SG, Zheng H, Yu B, Xie Z. Sevoflurane Acts on Ubiquitination-Proteasome Pathway to Reduce Postsynaptic Density 95 Protein Levels in Young Mice. Anesthesiology. 2017; 127:961–75. https://doi.org/10.1097/ALN.0000000000001889 [PubMed]
- 14. Yang Y, Zhong Z, Wang B, Xia X, Yao W, Huang L, Wang Y, Ding W. Early-life high-fat diet-induced obesity programs hippocampal development and cognitive functions via regulation of gut commensal Akkermansia muciniphila. Neuropsychopharmacology. 2019; 44:2054–64. https://doi.org/10.1038/s41386-019-0437-1 [PubMed]
- 15. Ji MH, Wang ZY, Sun XR, Tang H, Zhang H, Jia M, Qiu LL, Zhang GF, Peng YG, Yang JJ. Repeated Neonatal Sevoflurane Exposure-Induced Developmental Delays of Parvalbumin Interneurons and Cognitive Impairments Are Reversed by Environmental Enrichment. Mol Neurobiol. 2017; 54:3759–70. https://doi.org/10.1007/s12035-016-9943-x [PubMed]
- 16. Xue L, Zou X, Yang XQ, Peng F, Yu DK, Du JR. Chronic periodontitis induces microbiota-gut-brain axis disorders and cognitive impairment in mice. Exp Neurol. 2020; 326:113176. https://doi.org/10.1016/j.expneurol.2020.113176 [PubMed]
- 17. Liu G, Zhu T, Zhang A, Li F, Qian W, Qian B. Heightened stress response and cognitive impairment after repeated neonatal sevoflurane exposures might be linked to excessive GABAAR-mediated depolarization. J Anesth. 2016; 30:834–41. https://doi.org/10.1007/s00540-016-2215-0 [PubMed]
- 18. Zheng H, Dong Y, Xu Z, Crosby G, Culley DJ, Zhang Y, Xie Z. Sevoflurane anesthesia in pregnant mice induces neurotoxicity in fetal and offspring mice. Anesthesiology. 2013; 118:516–26. https://doi.org/10.1097/ALN.0b013e3182834d5d [PubMed]
- 19. Amrock LG, Starner ML, Murphy KL, Baxter MG. Long-term effects of single or multiple neonatal sevoflurane exposures on rat hippocampal ultrastructure. Anesthesiology. 2015; 122:87–95. https://doi.org/10.1097/ALN.0000000000000477 [PubMed]
- 20. Hofacer RD, Deng M, Ward CG, Joseph B, Hughes EA, Jiang C, Danzer SC, Loepke AW. Cell age-specific vulnerability of neurons to anesthetic toxicity. Ann Neurol. 2013; 73:695–704. https://doi.org/10.1002/ana.23892 [PubMed]
- 21. Culley DJ, Cotran EK, Karlsson E, Palanisamy A, Boyd JD, Xie Z, Crosby G. Isoflurane affects the cytoskeleton but not survival, proliferation, or synaptogenic properties of rat astrocytes in vitro. Br J Anaesth. 2013 (Suppl 1); 110:i19–28. https://doi.org/10.1093/bja/aet169 [PubMed]
- 22. O’Toole PW, Jeffery IB. Gut microbiota and aging. Science. 2015; 350:1214–15. https://doi.org/10.1126/science.aac8469 [PubMed]
- 23. Quigley EM. Microbiota-Brain-Gut Axis and Neurodegenerative Diseases. Curr Neurol Neurosci Rep. 2017; 17:94. https://doi.org/10.1007/s11910-017-0802-6 [PubMed]
- 24. Selkrig J, Wong P, Zhang X, Pettersson S. Metabolic tinkering by the gut microbiome: Implications for brain development and function. Gut Microbes. 2014; 5:369–80. https://doi.org/10.4161/gmic.28681 [PubMed]
- 25. Carabotti M, Scirocco A, Maselli MA, Severi C. The gut-brain axis: interactions between enteric microbiota, central and enteric nervous systems. Ann Gastroenterol. 2015; 28:203–09. [PubMed]
- 26. Cryan JF, O’Riordan KJ, Cowan CS, Sandhu KV, Bastiaanssen TF, Boehme M, Codagnone MG, Cussotto S, Fulling C, Golubeva AV, Guzzetta KE, Jaggar M, Long-Smith CM, et al. The Microbiota-Gut-Brain Axis. Physiol Rev. 2019; 99:1877–2013. https://doi.org/10.1152/physrev.00018.2018 [PubMed]
- 27. Bullich C, Keshavarzian A, Garssen J, Kraneveld A, Perez-Pardo P. Gut Vibes in Parkinson’s Disease: The Microbiota-Gut-Brain Axis. Mov Disord Clin Pract. 2019; 6:639–51. https://doi.org/10.1002/mdc3.12840 [PubMed]
- 28. Benakis C, Martin-Gallausiaux C, Trezzi JP, Melton P, Liesz A, Wilmes P. The microbiome-gut-brain axis in acute and chronic brain diseases. Curr Opin Neurobiol. 2020; 61:1–9. https://doi.org/10.1016/j.conb.2019.11.009 [PubMed]
- 29. Petra AI, Panagiotidou S, Hatziagelaki E, Stewart JM, Conti P, Theoharides TC. Gut-Microbiota-Brain Axis and Its Effect on Neuropsychiatric Disorders With Suspected Immune Dysregulation. Clin Ther. 2015; 37:984–95. https://doi.org/10.1016/j.clinthera.2015.04.002 [PubMed]
- 30. Obrenovich M, Jaworski H, Tadimalla T, Mistry A, Sykes L, Perry G, Bonomo RA. The Role of the Microbiota-Gut-Brain Axis and Antibiotics in ALS and Neurodegenerative Diseases. Microorganisms. 2020; 8:784. https://doi.org/10.3390/microorganisms8050784 [PubMed]
- 31. Serbanescu MA, Mathena RP, Xu J, Santiago-Rodriguez T, Hartsell TL, Cano RJ, Mintz CD. General Anesthesia Alters the Diversity and Composition of the Intestinal Microbiota in Mice. Anesth Analg. 2019; 129:e126–29. https://doi.org/10.1213/ANE.0000000000003938 [PubMed]
- 32. Li N, Wang Q, Wang Y, Sun A, Lin Y, Jin Y, Li X. Fecal microbiota transplantation from chronic unpredictable mild stress mice donors affects anxiety-like and depression-like behavior in recipient mice via the gut microbiota-inflammation-brain axis. Stress. 2019; 22:592–602. https://doi.org/10.1080/10253890.2019.1617267 [PubMed]
- 33. Ganci M, Suleyman E, Butt H, Ball M. The role of the brain-gut-microbiota axis in psychology: The importance of considering gut microbiota in the development, perpetuation, and treatment of psychological disorders. Brain Behav. 2019; 9:e01408. https://doi.org/10.1002/brb3.1408 [PubMed]
- 34. Yang M, Tan H, Zhang K, Lian N, Yu Y, Yu Y. Protective effects of Coenzyme Q10 against sevoflurane-induced cognitive impairment through regulating apolipoprotein E and phosphorylated Tau expression in young mice. Int J Dev Neurosci. 2020; 80:418–28. https://doi.org/10.1002/jdn.10041 [PubMed]
- 35. Liu Z, Dai X, Zhang H, Shi R, Hui Y, Jin X, Zhang W, Wang L, Wang Q, Wang D, Wang J, Tan X, Ren B, et al. Gut microbiota mediates intermittent-fasting alleviation of diabetes-induced cognitive impairment. Nat Commun. 2020; 11:855. https://doi.org/10.1038/s41467-020-14676-4 [PubMed]
- 36. Nadal A, Quesada I, Tudurí E, Nogueiras R, Alonso-Magdalena P. Endocrine-disrupting chemicals and the regulation of energy balance. Nat Rev Endocrinol. 2017; 13:536–46. https://doi.org/10.1038/nrendo.2017.51 [PubMed]
- 37. Yu F, Han W, Zhan G, Li S, Xiang S, Zhu B, Jiang X, Yang L, Luo A, Hua F, Yang C. Abnormal gut microbiota composition contributes to cognitive dysfunction in streptozotocin-induced diabetic mice. Aging (Albany NY). 2019; 11:3262–79. https://doi.org/10.18632/aging.101978 [PubMed]
- 38. Dimova LG, Zlatkov N, Verkade HJ, Uhlin BE, Tietge UJ. High-cholesterol diet does not alter gut microbiota composition in mice. Nutr Metab (Lond). 2017; 14:15. https://doi.org/10.1186/s12986-017-0170-x [PubMed]
- 39. Prager O, Friedman A, Nebenzahl YM. Role of neural barriers in the pathogenesis and outcome of Streptococcus pneumoniae meningitis. Exp Ther Med. 2017; 13:799–809. https://doi.org/10.3892/etm.2017.4082 [PubMed]
- 40. Watanabe I, Kuriyama N, Miyatani F, Nomura R, Naka S, Nakano K, Ihara M, Iwai K, Matsui D, Ozaki E, Koyama T, Nishigaki M, Yamamoto T, et al. Oral Cnm-positive Streptococcus Mutans Expressing Collagen Binding Activity is a Risk Factor for Cerebral Microbleeds and Cognitive Impairment. Sci Rep. 2016; 6:38561. https://doi.org/10.1038/srep38561 [PubMed]
- 41. Guo H, Chou WC, Lai Y, Liang K, Tam JW, Brickey WJ, Chen L, Montgomery ND, Li X, Bohannon LM, Sung AD, Chao NJ, Peled JU, et al. Multi-omics analyses of radiation survivors identify radioprotective microbes and metabolites. Science. 2020; 370:eaay9097. https://doi.org/10.1126/science.aay9097 [PubMed]
- 42. Barichella M, Severgnini M, Cilia R, Cassani E, Bolliri C, Caronni S, Ferri V, Cancello R, Ceccarani C, Faierman S, Pinelli G, De Bellis G, Zecca L, et al. Unraveling gut microbiota in Parkinson’s disease and atypical Parkinsonism. Mov Disord. 2019; 34:396–405. https://doi.org/10.1002/mds.27581 [PubMed]
- 43. Bajaj JS, Fagan A, White MB, Wade JB, Hylemon PB, Heuman DM, Fuchs M, John BV, Acharya C, Sikaroodi M, Gillevet PM. Specific Gut and Salivary Microbiota Patterns Are Linked With Different Cognitive Testing Strategies in Minimal Hepatic Encephalopathy. Am J Gastroenterol. 2019; 114:1080–90. https://doi.org/10.14309/ajg.0000000000000102 [PubMed]
- 44. Pérez-Burillo S, Pastoriza S, Fernández-Arteaga A, Luzón G, Jiménez-Hernández N, D’Auria G, Francino MP, Rufián-Henares JÁ. Spent Coffee Grounds Extract, Rich in Mannooligosaccharides, Promotes a Healthier Gut Microbial Community in a Dose-Dependent Manner. J Agric Food Chem. 2019; 67:2500–09. https://doi.org/10.1021/acs.jafc.8b06604 [PubMed]
- 45. Louis S, Tappu RM, Damms-Machado A, Huson DH, Bischoff SC. Characterization of the Gut Microbial Community of Obese Patients Following a Weight-Loss Intervention Using Whole Metagenome Shotgun Sequencing. PLoS One. 2016; 11:e0149564. https://doi.org/10.1371/journal.pone.0149564 [PubMed]
- 46. Zhan G, Yang N, Li S, Huang N, Fang X, Zhang J, Zhu B, Yang L, Yang C, Luo A. Abnormal gut microbiota composition contributes to cognitive dysfunction in SAMP8 mice. Aging (Albany NY). 2018; 10:1257–67. https://doi.org/10.18632/aging.101464 [PubMed]
- 47. Sturman DA, Moghaddam B. The neurobiology of adolescence: changes in brain architecture, functional dynamics, and behavioral tendencies. Neurosci Biobehav Rev. 2011; 35:1704–12. https://doi.org/10.1016/j.neubiorev.2011.04.003 [PubMed]
- 48. Ing C, DiMaggio C, Whitehouse A, Hegarty MK, Brady J, von Ungern-Sternberg BS, Davidson A, Wood AJ, Li G, Sun LS. Long-term differences in language and cognitive function after childhood exposure to anesthesia. Pediatrics. 2012; 130:e476–85. https://doi.org/10.1542/peds.2011-3822 [PubMed]
- 49. Li Y, Liu C, Zhao Y, Hu K, Zhang J, Zeng M, Luo T, Jiang W, Wang H. Sevoflurane induces short-term changes in proteins in the cerebral cortices of developing rats. Acta Anaesthesiol Scand. 2013; 57:380–90. https://doi.org/10.1111/aas.12018 [PubMed]
- 50. Burnett S, Sebastian C, Cohen Kadosh K, Blakemore SJ. The social brain in adolescence: evidence from functional magnetic resonance imaging and behavioural studies. Neurosci Biobehav Rev. 2011; 35:1654–64. https://doi.org/10.1016/j.neubiorev.2010.10.011 [PubMed]
- 51. Desbonnet L, Clarke G, Traplin A, O’Sullivan O, Crispie F, Moloney RD, Cotter PD, Dinan TG, Cryan JF. Gut microbiota depletion from early adolescence in mice: Implications for brain and behaviour. Brain Behav Immun. 2015; 48:165–73. https://doi.org/10.1016/j.bbi.2015.04.004 [PubMed]
- 52. Song SY, Meng XW, Xia Z, Liu H, Zhang J, Chen QC, Liu HY, Ji FH, Peng K. Cognitive impairment and transcriptomic profile in hippocampus of young mice after multiple neonatal exposures to sevoflurane. Aging (Albany NY). 2019; 11:8386–417. https://doi.org/10.18632/aging.102326 [PubMed]
- 53. Shen X, Dong Y, Xu Z, Wang H, Miao C, Soriano SG, Sun D, Baxter MG, Zhang Y, Xie Z. Selective anesthesia-induced neuroinflammation in developing mouse brain and cognitive impairment. Anesthesiology. 2013; 118:502–15. https://doi.org/10.1097/ALN.0b013e3182834d77 [PubMed]
- 54. Farzi A, Fröhlich EE, Holzer P. Gut Microbiota and the Neuroendocrine System. Neurotherapeutics. 2018; 15:5–22. https://doi.org/10.1007/s13311-017-0600-5 [PubMed]
- 55. Parker A, Lawson MA, Vaux L, Pin C. Host-microbe interaction in the gastrointestinal tract. Environ Microbiol. 2018; 20:2337–53. https://doi.org/10.1111/1462-2920.13926 [PubMed]
- 56. Hernández-Chirlaque C, Aranda CJ, Ocón B, Capitán-Cañadas F, Ortega-González M, Carrero JJ, Suárez MD, Zarzuelo A, Sánchez de Medina F, Martínez-Augustin O. Germ-free and Antibiotic-treated Mice are Highly Susceptible to Epithelial Injury in DSS Colitis. J Crohns Colitis. 2016; 10:1324–35. https://doi.org/10.1093/ecco-jcc/jjw096 [PubMed]
- 57. Liufu N, Liu L, Shen S, Jiang Z, Dong Y, Wang Y, Culley D, Crosby G, Cao M, Shen Y, Marcantonio E, Xie Z, Zhang Y. Anesthesia and surgery induce age-dependent changes in behaviors and microbiota. Aging (Albany NY). 2020; 12:1965–86. https://doi.org/10.18632/aging.102736 [PubMed]
- 58. de Vos WM, de Vos EA. Role of the intestinal microbiome in health and disease: from correlation to causation. Nutr Rev. 2012 (Suppl 1); 70:S45–56. https://doi.org/10.1111/j.1753-4887.2012.00505.x [PubMed]
- 59. Lozupone CA, Stombaugh JI, Gordon JI, Jansson JK, Knight R. Diversity, stability and resilience of the human gut microbiota. Nature. 2012; 489:220–30. https://doi.org/10.1038/nature11550 [PubMed]
- 60. Adlerberth I, Wold AE. Establishment of the gut microbiota in Western infants. Acta Paediatr. 2009; 98:229–38. https://doi.org/10.1111/j.1651-2227.2008.01060.x [PubMed]
- 61. Marques TM, Wall R, Ross RP, Fitzgerald GF, Ryan CA, Stanton C. Programming infant gut microbiota: influence of dietary and environmental factors. Curr Opin Biotechnol. 2010; 21:149–56. https://doi.org/10.1016/j.copbio.2010.03.020 [PubMed]
- 62. Koenig JE, Spor A, Scalfone N, Fricker AD, Stombaugh J, Knight R, Angenent LT, Ley RE. Succession of microbial consortia in the developing infant gut microbiome. Proc Natl Acad Sci USA. 2011 (Suppl 1); 108:4578–85. https://doi.org/10.1073/pnas.1000081107 [PubMed]
- 63. Borre YE, Moloney RD, Clarke G, Dinan TG, Cryan JF. The impact of microbiota on brain and behavior: mechanisms and therapeutic potential. Adv Exp Med Biol. 2014; 817:373–403. https://doi.org/10.1007/978-1-4939-0897-4_17 [PubMed]
- 64. Cryan JF, Dinan TG. Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nat Rev Neurosci. 2012; 13:701–12. https://doi.org/10.1038/nrn3346 [PubMed]
- 65. Falony G, Vieira-Silva S, Raes J. Richness and ecosystem development across faecal snapshots of the gut microbiota. Nat Microbiol. 2018; 3:526–28. https://doi.org/10.1038/s41564-018-0143-5 [PubMed]
- 66. Eckburg PB, Bik EM, Bernstein CN, Purdom E, Dethlefsen L, Sargent M, Gill SR, Nelson KE, Relman DA. Diversity of the human intestinal microbial flora. Science. 2005; 308:1635–38. https://doi.org/10.1126/science.1110591 [PubMed]
- 67. Satomoto M, Satoh Y, Terui K, Miyao H, Takishima K, Ito M, Imaki J. Neonatal exposure to sevoflurane induces abnormal social behaviors and deficits in fear conditioning in mice. Anesthesiology. 2009; 110:628–37. https://doi.org/10.1097/ALN.0b013e3181974fa2 [PubMed]
- 68. Tao G, Zhang J, Zhang L, Dong Y, Yu B, Crosby G, Culley DJ, Zhang Y, Xie Z. Sevoflurane induces tau phosphorylation and glycogen synthase kinase 3β activation in young mice. Anesthesiology. 2014; 121:510–27. https://doi.org/10.1097/ALN.0000000000000278 [PubMed]
- 69. Zhong JY, Magnusson KR, Swarts ME, Clendinen CA, Reynolds NC, Moffat SD. The application of a rodent-based Morris water maze (MWM) protocol to an investigation of age-related differences in human spatial learning. Behav Neurosci. 2017; 131:470–82. https://doi.org/10.1037/bne0000219 [PubMed]
- 70. Ge X, Zhao W, Ding C, Tian H, Xu L, Wang H, Ni L, Jiang J, Gong J, Zhu W, Zhu M, Li N. Potential role of fecal microbiota from patients with slow transit constipation in the regulation of gastrointestinal motility. Sci Rep. 2017; 7:441. https://doi.org/10.1038/s41598-017-00612-y [PubMed]
- 71. Yang C, Fujita Y, Ren Q, Ma M, Dong C, Hashimoto K. Bifidobacterium in the gut microbiota confer resilience to chronic social defeat stress in mice. Sci Rep. 2017; 7:45942. https://doi.org/10.1038/srep45942 [PubMed]
- 72. Magoč T, Salzberg SL. FLASH: fast length adjustment of short reads to improve genome assemblies. Bioinformatics. 2011; 27:2957–63. https://doi.org/10.1093/bioinformatics/btr507 [PubMed]
- 73. Yoon SH, Ha SM, Kwon S, Lim J, Kim Y, Seo H, Chun J. Introducing EzBioCloud: a taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int J Syst Evol Microbiol. 2017; 67:1613–17. https://doi.org/10.1099/ijsem.0.001755 [PubMed]
- 74. Segata N, Izard J, Waldron L, Gevers D, Miropolsky L, Garrett WS, Huttenhower C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011; 12:R60. https://doi.org/10.1186/gb-2011-12-6-r60 [PubMed]