



Introduction
Bronchial asthma is characterized by airway hyperresponsiveness (AHR), variable airflow limitation, and airway remodeling [1]. Currently, there is no effective therapy for this disease [1, 2]. Therefore, it is critical to determine the molecular mechanisms of asthma.
Although the airway epithelial cells serve as a first defending barrier of defense against invading pathogens in asthma, recent studies have indicated that macrophages play an important role in fighting against pathogenic invasion [3, 4]. Bang et al. demonstrated that allergic airway inflammation was significantly ameliorated when unsensitized alveolar macrophages (AMs) were adoptively transferred to macrophage-depleted sensitized mice [5]. However, the underlying mechanisms of AMs in regulation of allergic airway inflammatory remain unknown.
Sirtuin 1 (SIRT1) plays an important role in regulating many pathophysiological processes, such as inflammation, autoimmunity, and apoptosis [6]. Our previous study has shown that serum SIRT1 levels in patients with asthma were positively associated serum IgE levels [7]. Ichikawa et al. found that SIRT1 activator suppressed inflammatory cell infiltration and cytokine production in asthmatic mice [8]. Kardan et al. discovered that SIRT1 protein levels were decreased in patients with severe asthma [4]. Although AMs are the first responders to foreign pathogens in the airway and secrete proinflammatory response upon exposure to outside invaders, the possible role of SIRT1 in airway inflammation of macrophages following allergen exposure remains unclear.
Here, using an experimental model of asthma, we sought to determine the underlying mechanisms of SIRT1 in the regulation of allergic airway inflammatory responses in macrophages using an experimental model of asthma. Our findings revealed that ERK/p38 MAPK pathways being the underlying mechanism by which SIRT1 regulates the response of macrophages upon allergen exposure to regulate airway inflammation and mucus production.
Results
SIRT1 expression is decreased in macrophages following allergen exposure
We first assessed SIRT1expression in the lung tissue of mice. Allergic asthma model was established in of Sirt1fl/fl mice, as described in Figure 1A. The results IHC staining revealed that the expression of SIRT1 was significantly decreased in lung tissue of asthmatic Sirt1fl/fl mice compared with those in control groups (Figure 1B, 1C). The expression of SIRT1 in the lung tissue of model mice was further confirmed by WB and RT-PCR (Figure 1D, 1E). We next sought to identify specific cell types related to abnormal expression of SIRT1. We found that SIRT1 was mainly expressed in lung macrophages of asthmatic Sirt1fl/fl mice (Figure 1F, 1G), and similar results were found in the bronchoalveolar lavage (BALF) samples (Figure 1H, 1I).
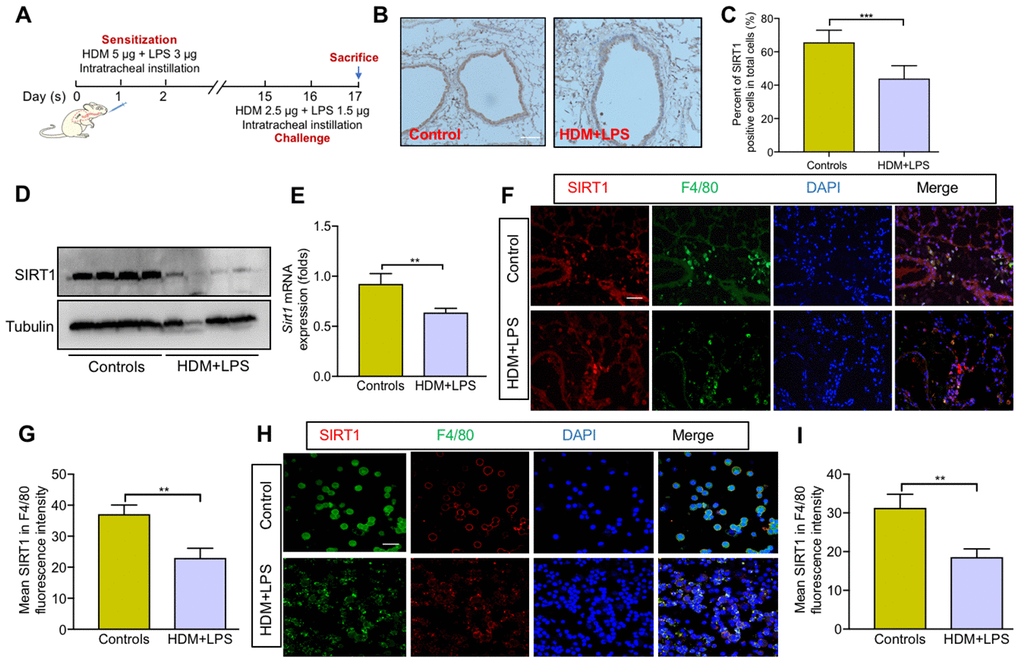
Figure 1. SIRT1 expression is decreased in macrophages following allergen exposure. (A) Schematic map of allergic asthma model; (B) Representative IHC images of SIRT1 in lung tissue of mice (scale bar, 100 μm); (C) Semiquantification of IHC was done using Image Pro 6.1 software; (D, E) SIRT1 expression in lung tissues of Sirt1fl/fl mice were assessed using Western blot and RT-PCR analysis; Representative immunofluorescence images of SIRT1 expression in alveolar macrophages are revealed using F4/80 in lung tissues (F) and BALF (H) of mice (Scale bar, 100 μm and 20 μm, respectively); (G, I) Semiquantification of immunofluorescence images was done using Image Pro 6.1 software. Data are presented as Mean ± SEM of three independent experiments (n = 5-8 for each group). **P<0.01 and ***P<0.001.
To further explore the role of SIRT1 in lung macrophages, we generated BMDMs from Sirt1fl/fl mice. Consistent with the above findings, the expression of Sirt1 gene was significantly reduced in HDM-induced BMDMs in a dose- and time-dependent manner (Figure 2A, 2B). SIRT1 protein levels in BMDMs were confirmed by using WB and immunofluorescence (IF) analysis (Figure 2C–2E). Taken together, these findings suggested that macrophages are involved in SIRT1-mediated airway allergic inflammation.
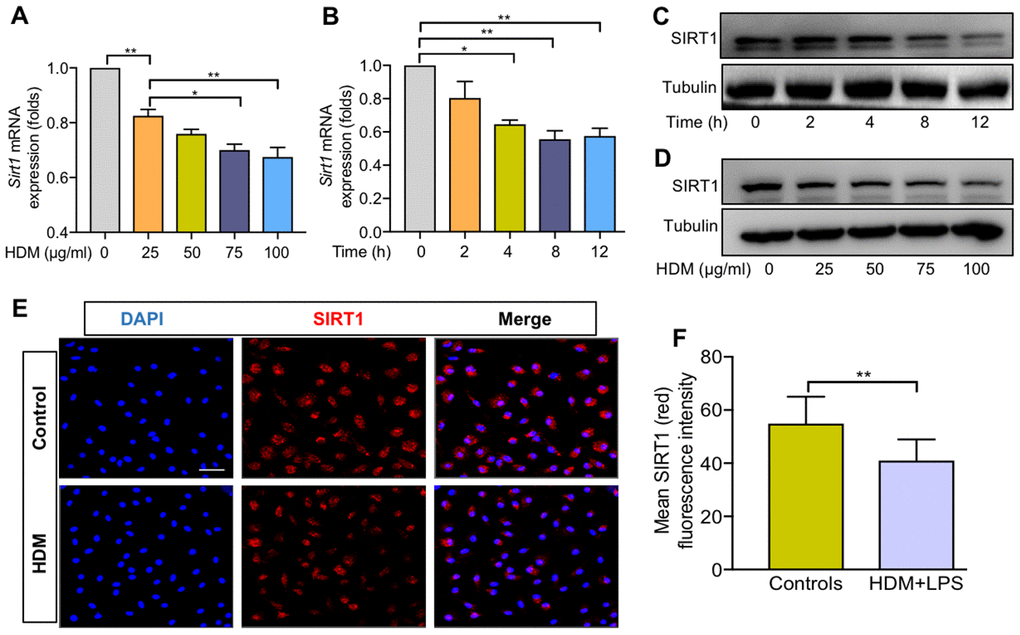
Figure 2. SIRT1 expression was decreased in BMDMs exposed to HDM. Dose–dependent expression of SIRT1 in bone marrow–derived macrophages (BMDMs) stimulated with HDM for 24 h and was analyzed using RT-PCR (A) and WB (C); Time–dependent expression of SIRT1 in BMDMs treated with HDM at 100 μg/ml and was analyzed using RT-PCR (B) and Western blot (D); BMDMs treated with HDM at 100 μg/ml for 24 h. Representative immunofluorescence images of SIRT1 (red) expression in BMDMs (E, Scale bar, 50 μm) and semiquantification of immunofluorescence images was done using Image Pro 6.1 software (F). Data are presented as Mean ± SEM of three independent experiments. *P<0.05, **P<0.01 and ***P<0.001.
Sirt1-deficient BMDMs increase cytokine secretion following HDM exposure
We next generated mice with myeloid conditional deletion of Sirt1 (mainly in macrophages) (Sirt1fl/fl-LysMcre, Figure 3A). Representative genotyping results as shown in Figure 3B. The genotyping results were further determined by WB and RT-PCR analysis (Figure 3C, 3D). Sirt1fl/fl-LysMCre mice had a normal weight and lifespan. No pathological changes were observed in the lungs of Sirt1fl/fl-LysMCre mice (Supplementary Figure 1A). Moreover, Sirt1fl/fl-LysMCre mice did not show defects in the development and differentiation of immune cells (Supplementary Figure 1B–1F).
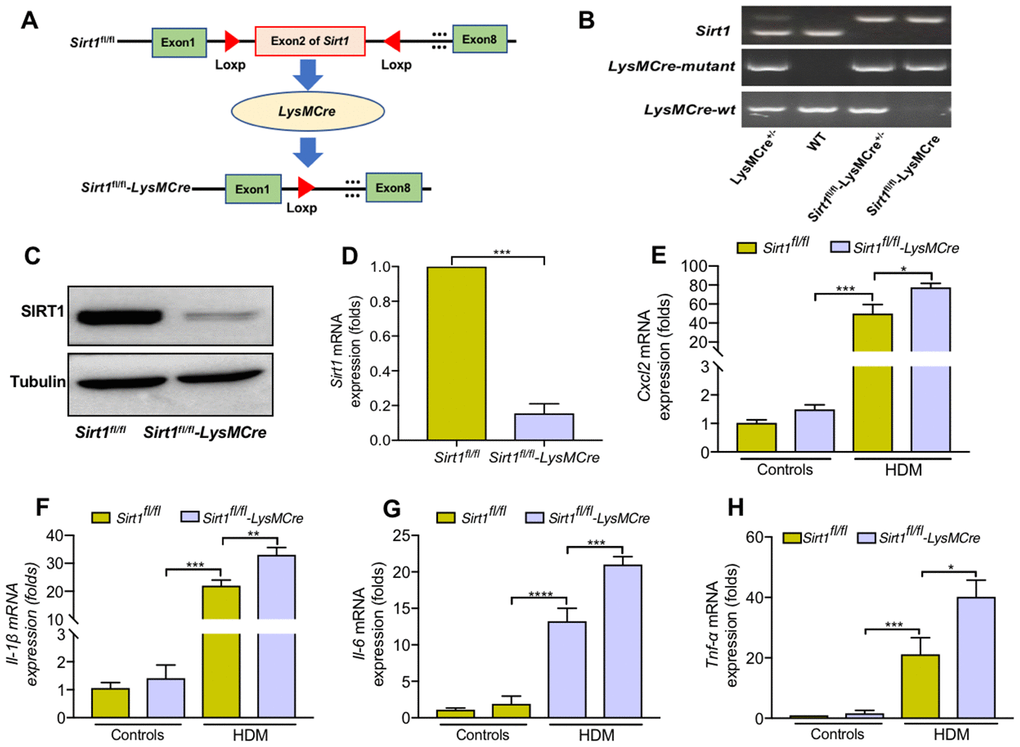
Figure 3. Sirt1-deficient BMDMs enhanced cytokine secretion when treated with HDM. (A) Schematic map of the generation of Sirt1fl/fl-LysMCre mice. (B) Genotyping was performed by PCR using mouse tail genomic DNA. (C, D) Genotyping was assessed in BMDMs from Sirt1fl/fl and Sirt1fl/fl-LysMCre mice using Western blot and RT-PCR analysis. (E–H) BMDMs were treated with HDM at 100 μg/ml for 24 h to measure the levels of Cxcl2, Il-1β, Il-6, and Tnf-α. Data are presented as Mean ± SEM of three independent experiments. *P<0.05, **P<0.01, ***P<0.001 and ****P<0.0001.
We generated BMDMs from Sirt1fl/fl-LysMCre and Sirt1fl/fl mice, and found that Il-6, Tnf-α, Cxcl2, and Il-1β expression were significantly increased in Sirt1-deficient BMDMs compared with the control group (Figure 3E–3H). Collectively, these findings imply that loss of SIRT1 in macrophages enhanced inflammatory cytokine response.
Sirt1fl/fl-LysMCre mice display exacerbated airway inflammation
To further elucidate the role of SIRT1 in allergic airway inflammation in vivo, we established allergic asthma model using Sirt1fl/fl littermates and Sirt1fl/fl-LysMCre mice, as described in Figure 1A Compared with asthmatic Sirt1fl/fl mice, SIRT1 expression in lung tissue was significantly decreased in asthmatic Sirt1fl/fl-LysMCre mice (Figure 4A).
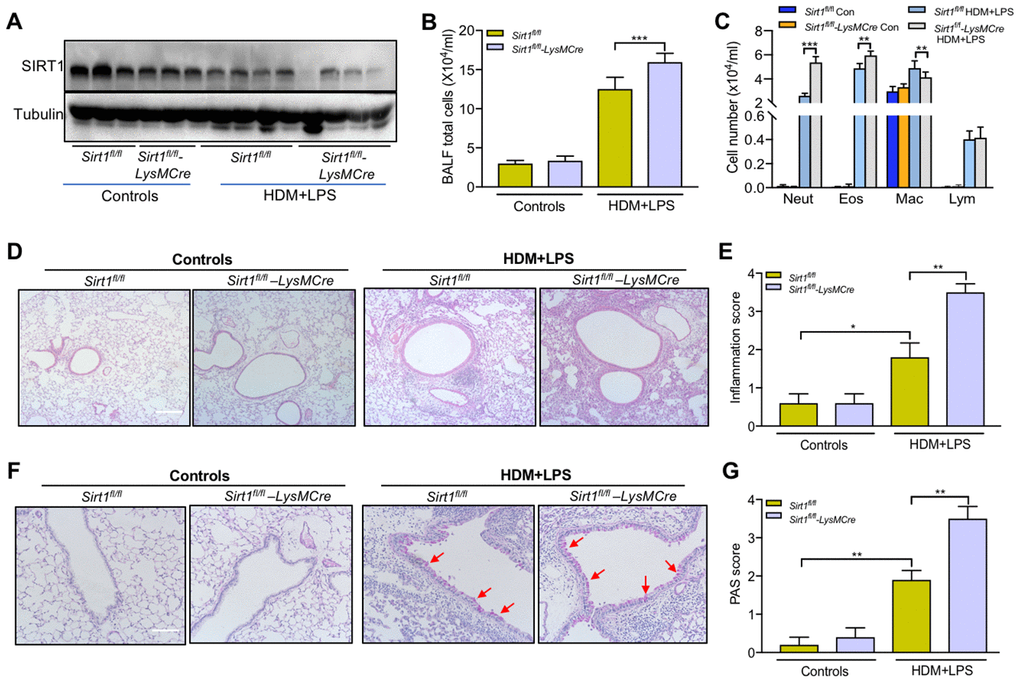
Figure 4. Impairing of SIRT1 exacerbates airway inflammation following allergen exposure.Sirt1fl/fl-LysMCre mice and Sirt1fl/fl littermates were established allergic asthma model as described in the methods section (n = 5-8 for each group). (A) SIRT1 expression in lung tissues of Sirt1fl/fl and Sirt1fl/fl-LysMCre mice were assessed using Western blot; (B, C) Total cells and differential cell counts in BALF were measured; (D) Representative photomicrographs of lung inflammation expression are shown (Scale bar, 100 μm); (E) Semiquantification of inflammation expression in the lungs were preformed using Image Pro 6.1 software; (F) Representative photomicrographs of mucus production are shown (red arrows) (Scale bar, 100 μm); (G) Semiquantification of mucus production in the lungs were preformed using Image Pro 6.1 software. Data are presented as Mean ± SEM of three independent experiments. *P<0.05, **P<0.01 and ***P<0.001.
The total BALF cells, eosinophils and neutrophils were significantly increased in BALF of asthmatic Sirt1fl/fl-LysMCre mice compared with those in asthmatic Sirt1fl/fl mice (Figure 4B, 4C). Moreover, airway inflammation and mucus secretion were significantly increased in Sirt1fl/fl-LysMCre mice exposed to the allergen (Figure 4D–4G). Compared with the Sirt1fl/fl mice, both mRNA and protein expression of TNF-α, CXCL2, and IL-1β in the lung tissue were significantly enhanced in allergen-exposed Sirt1fl/fl-LysMCre mice (Figure 5).
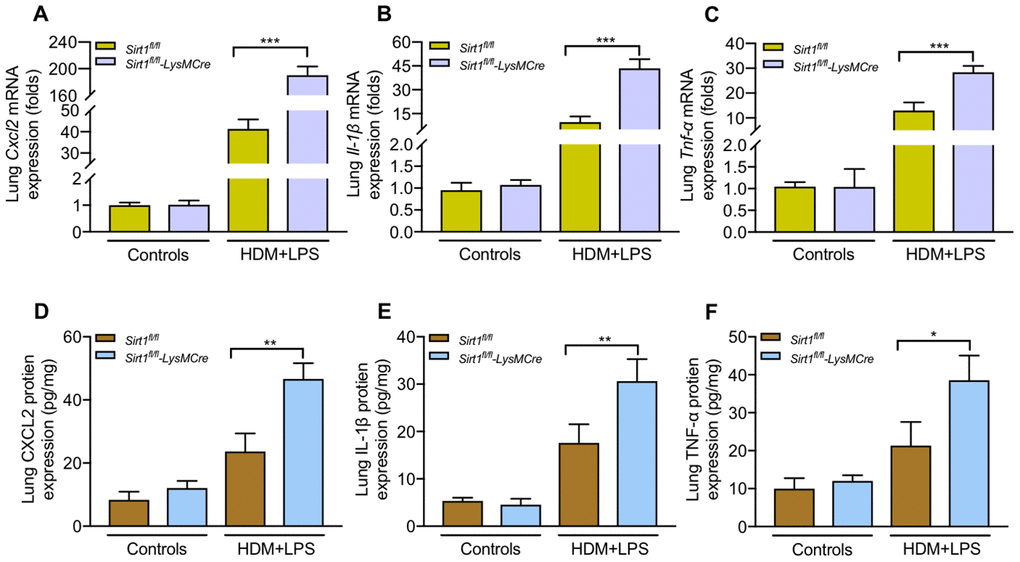
Figure 5. Sirt1-deficient exacerbates inflammatory cytokines production following allergen exposure.Sirt1fl/fl-LysMCre mice and Sirt1fl/fl littermates were established allergic asthma model (n = 5-8 mice per group per study). Lungs were isolated 1 day after the last challenge. (A–C) Expression of the mRNA levels of inflammatory cytokines in the lung homogenate were analyzed by RT-PCR; (D–F) Protein levels of inflammatory cytokines in the lung homogenate were measured by ELISA. Data are representative of three independent experiments with similar results. Data are presented as Mean ± SEM of three independent experiments. *P<0.05, **P<0.01 and ***P<0.001.
SIRT1 suppresses airway inflammation partially through inhibition of ERK/p38 MAPK pathway activation
Finally, we assessed the underlying pathways that mediate SIRT1 in regulation of airway inflammatory in macrophages. The mitogen-activated protein kinase (MAPK) pathways is involved in airway inflammation. Therefore, we sought to determine whether MAPK pathways mediate the role of SIRT1 in regulation of inflammatory responses. We found that the phosphorylation of p38 and ERK was increased in HDM-induced BMDMs (Supplementary Figure 2). Moreover, the p-p38 and p-ERK expression in Sirt1fl/fl-LysMCre mice were further aggravated compared with the Sirt1fl/fl mice (Figure 6A, 6B).
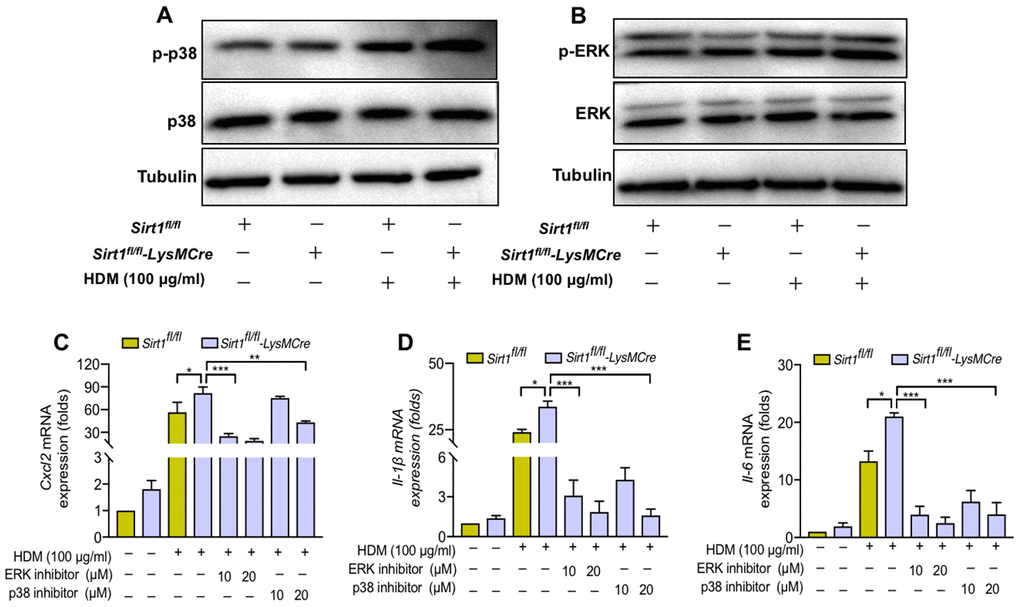
Figure 6. SIRT1 suppresses airway inflammation partially through inhibition of ERK/p38 MAPK pathway activation. (A, B) Sirt1fl/fl and Sirt1fl/fl-LysMCre BMDMs were treated with HDM, the expression of p-p38 and p-ERK was assessed by using Western blot analysis; (C–E) p38 inhibitor SB203580 and ERK inhibitor U0126 were added in HDM-exposed BMDMs and BMDMs were collected after 24 h. The levels of Cxcl2, Il-1β, and Il6 were analyzed by RT-PCR. Data are representative of three independent studies. Results are expressed as mean ± SEM. *P<0.05, **P<0.01 and ***P<0.001.
To further validate the role of ERK/p38 MAPK pathway in SIRT1 mediated- airway inflammation in asthma, we used the p38 inhibitor SB203580 and the ERK inhibitor U0126 to examine whether blocking ERK/p38 MAPK activation could reverse the amplified cytokine production caused by SIRT1 deficiency. We found that the levels of inflammatory cytokines were significantly decreased in SIRT1-deficient BMDMs treated with SB203580 and U0126. (Figure 6C–6E).
These results suggested that SIRT1 deficiency enhanced ERK/p38 activation and cytokine production in macrophages.
Discussion
The present study showed that SIRT1 expression was decreased in HDM-induced BMDMs while Sirt1-deficient BMDMs increase cytokine secretion when treated with HDM. Moreover, mice with myeloid cells specific knockdown of SIRT1, display exacerbated airway inflammation. ERK/p38 MAPK pathway is the possible underlying mechanism of SIRT1 mediated suppression of HDM-induced airway inflammation in macrophages. Our study suggested that activation of SIRT1 in macrophages may represent therapeutic strategy for asthma.
Sirtuins regulate diverse biological functions such as stress resistance, metabolism, and aging. SIRT1 plays a critical role in inflammatory diseases by regulating transcription factors, airway inflammation, apoptosis and senescence [9]. However, the roles of SIRT1 in regulating macrophage activation in the pathogenesis of asthma remain unclear [5]. Macrophages from healthy subjects produce lesser IL-10 and co-stimulatory molecules (CD80 and CD86) than asthmatic patients [10]. A study revealed that macrophages from unsensitized mice provided protection against asthmatic symptoms [11]. In contrast, other studies demonstrated that macrophages play a minor role in airway allergic inflammation [12, 13]. The disparity in these studies may be attributed to the different macrophage studied. Our study focused on the function of macrophages in regulation of HDM-induced airway inflammation.
SIRT1 is involved in regulation of the inflammatory response in macrophages [14, 15]. It has been reported that SIRT1 induced macrophage polarization and attenuated monosodium urate crystal-induced inflammation [16]. Myeloid-specific deletion of SIRT1 enhanced obesity-induced inflammation [17]. Similarly, mouse macrophage specific knockout of SIRT1 exacerbated abdominal aortic aneurysm formation [18]. In the context of insulin resistance, macrophage SIRT1 may play a beneficial role in regulating glucose homeostasis [19]. However, the mechanisms of macrophage SIRT1 in airway allergic inflammation remain largely unknown. Here, we found that SIRT1 expression is reduced in HDM-induced Sirt1fl/fl BMDMs. HDM exposure significantly enhanced cytokine production in Sirt1fl/fl BMDMs, and BMDMs deficient in Sirt1 further increased cytokine production. Consistent with in vitro experiments, Sirt1fl/fl-LysMCre mice showed substantially enhanced airway inflammation, cytokine production, and mucus production compared with allergen exposed Sirt1fl/fl-LysMCre mice. Further, we found that SIRT1 suppresses airway inflammation at least partially through inhibition of ERK/p38 MAPK pathway activation. Collectively, these data suggest that downregulation of SIRT1 could be a necessary mechanism by which allergic airway inflammation occurs.
Our study does have some limitations. First, macrophages are polarized towards the M2 phenotype in asthma. Whether SIRT1 mediates the M2-type AM polarization and involves in the pathogenesis of asthma remain to be further elucidated. Second, since SIRT1 is an NAD-dependent deacetylase, SIRT1's deacetylation activity via epigenetics is one of its most important functions. Thus, the epigenetic role of SIRT1 in macrophages of asthma also needs to be further investigated.
In conclusion, our study reveals that SIRT1 suppresses the allergic airway inflammation in macrophages, and suggested that activation of SIRT1 in macrophages might be a strategy for treating the allergic airway inflammation.
Materials and Methods
Animal studies
The LysMcre mice were a generous gift from Dr. G. Feng (University of California at San Diego, CA, USA). Sirt1fl/fl mice were purchased from GemPharmatech Co., Ltd., and. Myeloid cell-specific SIRT1conditional knockout mice (Sirt1fl/fl-LysMCre) were obtained by crossing Sirt1fl/fl mice with the LysMCre mice. Sequence of primers for LysMCre and Sirt1 genes are listed in Table 1. All experimental procedures were approved by the Guangdong Medical University’s Animal Ethical Committee.
Table 1. Sequence of primers were used in this study.
| Genes | Forward | Reverse |
| Sirt1 (m) | 5ʹ-CACTGTAACTGGGGGCAACT-3’ | 5ʹ-CACTTCTTGTCAGCGTCGAA-3’ |
| Cxcl-2(m) | 5ʹ-TGTCCCTCAACGGAAGAACC-3’ | 5ʹ-CTCAGACAGCGAGGCACATC-3’ |
| Il-1β(m) | 5ʹ-GCAACTGTTCCTGAACTCAACT-3’ | 5ʹ-ATCTTTTGGGGTCCGTCAACT-3’ |
| Tnf-α(m) | 5ʹ-GACGTGGAACTGGCAGAAGAG-3’ | 5ʹ-TTGGTGGTTTGTGAGTGTGAG-3’ |
| β-actin(m) | 5ʹ-AGTGTGACGTTGACATCCGT-3’ | 5ʹ-GCAGCTCAGTAACAGTCCGC-3’ |
| LysMCre-WT(m) | 5ʹ-TTACAGTCGGCCAGGCTGAC-3’ | |
| LysMCre-Mutant(m) | 5ʹ-CCCAG AAATGCCAGATTACG-3’ | |
| LysMCre-Common(m) | 5ʹ-CTTGGGCTGCCAGAATTTCTC-3’ | |
| Sirt1 gene deletion(m) | oIMR7909: 5ʹ-GGTTGACTTAGGTCTTGTCTG-3’ | |
| oIMR7912: 5ʹ-CGTCCCTTGTAATGTTTCCC-3’ | ||
BMDMs
BMDMs were isolated and cultured as described previously [20]. Briefly, BM cells were collected from six- to eight week-old male mice. Red blood cells (RBCs) Lysing Buffer (Tbdscience.com) was used to remove red blood cells. The remaining cells were incubated in DMEM containing antibiotics, FBS (10%), and 10 ng/ml recombinant mouse M-CSF (Novoprotein, Catalog #0331488) for 7 days to promote differentiate bone marrow-derived macrophages.
Allergic asthma mouse model
Allergic asthma mouse model was established according to previous studies [21–23]. Briefly, six- to eight-week-old mice were sensitized by intratracheal instillation of 5 μg HDM (Greer Laboratories) plus 3 μg LPS (Sigma) in 50 μl normal saline (NS) or with 50 μl NS on days 0, 1, and 2. On days 15 and 16 after the initial sensitization, the mice were challenged with 2.5 μg HDM plus 1.5 μg LPS in 50 μl NS or with 50 μl NS using intratracheal instillation. The mice were sacrificed under anesthesia 24 h after the last challenge. Bronchoalveolar lavage fluid (BALF) was conducted according to our previous study [2]. Cytospin slides were performed using the Wright-Giemsa staining method. Hematoxylin/eosin (HE) staining sections and mucus production of goblet cells were assessed using a semiquantitative scoring system as previously described [17]. RT-PCR analysis was preformed according to our previous study [2]. The sequence of primers is listed in Table 1.
Immunohistochemistry (IHC)
The slides were detected with a rabbit polyclonal antibody against SIRT1 (Abcam). Semiquantitative evaluation of SIRT1 positive cells was analyzed with the Image Pro 6.1 software following previously described methods [2].
Western blot
Lysates from BMDMs and lung tissues were separated by SDS-PAGE and immunoblotted as previously described methods [2], using antibodies against the following proteins: SIRT1 (Abcam), Tubuin (Beyotime), p-ERK1/2, ERK, p38, and p-p38 (CST).
ELISA
The expression of TNF-α, CXCL2, and IL-1β in lung homogenate were detected by ELISA kits (eBioscience) following the manufacturer’s recommendation.
Flow cytometry
Flow cytometry was performed on a Fortessa (BD Biosciences) as previously described methods [2]. Anti-mouse CD19-Percp, CD8-PE-Cy7, F4/80-APC, CD3-APC-Cy7, CD11c-PE, CD4-FITC, and CD49b-PE were purchased from Biolegend.
Immunofluorescence (IF)
The expression of SIRT1 in alveolar macrophages (AMs) and BMDM cells was detected by immunofluorescence as previously described methods [2]. DAPI was used for nucleus staining.
Statistical analysis
Results are described as mean with SEM. All calculations and the graph were conducted using GraphPad Prism (San Diego, CA, version 8.0). Mann-Whitney U-test or Student's t test was used to compare differences between two groups. For comparisons between more than two groups, one-way ANOVA was applied. A value of P less than 0.05 was considered statistically significant.
Supplementary Materials
Author Contributions
T.L. and B.W. designed the experiments; T.L., G.S., Z.C. and Y.G. performed the experiments; Y.C., M.Z., H.Y., and C.C. collected clinical samples; T.L. and G.S. analyzed the data; T.L. and G.S. wrote the manuscript; B.W., T.L., D.W., M.C. and D.L. interpreted the results and revised the manuscript. All authors read and approved the final manuscript.
Acknowledgments
We thank Prof. Gensheng Feng (University of California at San Diego, CA) for providing the LysMCre mice. We thank Madiha Zahra Syeda (Zhejiang University) for correcting grammar errors on the manuscript.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Funding
This work was supported by the National Natural Science Foundation of China (81873404, 81670025), Guangdong Basic and Applied Basic Research Foundation (2020B1515020004, 2018A0303130269), Medical Scientific Research Foundation of Guangdong Province (No. A2018162). The Affiliated Hospital of Guangdong Medical University Clinical Research Program (LCYJ2018C001).
References
- 1. Lai T, Wu M, Zhang C, Che L, Xu F, Wang Y, Wu Y, Xuan N, Cao C, Du X, Wu B, Li W, Ying S, et al. HDAC2 attenuates airway inflammation by suppressing IL-17A production in HDM-challenged mice. Am J Physiol Lung Cell Mol Physiol. 2019; 316:L269–79. https://doi.org/10.1152/ajplung.00143.2018 [PubMed]
- 2. Global Initiative for Asthma. Global Strategy for Asthma Management and Prevention. 2019. https://ginasthma.org.
- 3. Balhara J, Gounni AS. The alveolar macrophages in asthma: a double-edged sword. Mucosal Immunol. 2012; 5:605–09. https://doi.org/10.1038/mi.2012.74 [PubMed]
- 4. Colley T, Mercado N, Kunori Y, Brightling C, Bhavsar PK, Barnes PJ, Ito K. Defective sirtuin-1 increases IL-4 expression through acetylation of GATA-3 in patients with severe asthma. J Allergy Clin Immunol. 2016; 137:1595–1597.e7. https://doi.org/10.1016/j.jaci.2015.10.013 [PubMed]
- 5. Bang BR, Chun E, Shim EJ, Lee HS, Lee SY, Cho SH, Min KU, Kim YY, Park HW. Alveolar macrophages modulate allergic inflammation in a murine model of asthma. Exp Mol Med. 2011; 43:275–80. https://doi.org/10.3858/emm.2011.43.5.028 [PubMed]
- 6. Noh SJ, Baek HA, Park HS, Jang KY, Moon WS, Kang MJ, Lee DG, Kim MH, Lee JH, Chung MJ. Expression of SIRT1 and cortactin is associated with progression of non-small cell lung cancer. Pathol Res Pract. 2013; 209:365–70. https://doi.org/10.1016/j.prp.2013.03.011 [PubMed]
- 7. Wang Y, Li D, Ma G, Li W, Wu J, Lai T, Huang D, Zhao X, Lv Q, Chen M, Wu B. Increases in peripheral SIRT1: a new biological characteristic of asthma. Respirology. 2015; 20:1066–72. https://doi.org/10.1111/resp.12558 [PubMed]
- 8. Ichikawa T, Hayashi R, Suzuki K, Imanishi S, Kambara K, Okazawa S, Inomata M, Yamada T, Yamazaki Y, Koshimizu Y, Miwa T, Matsui S, Usui I, et al. Sirtuin 1 activator SRT1720 suppresses inflammation in an ovalbumin-induced mouse model of asthma. Respirology. 2013; 18:332–39. https://doi.org/10.1111/j.1440-1843.2012.02284.x [PubMed]
- 9. Rahman I, Kinnula VL, Gorbunova V, Yao H. SIRT1 as a therapeutic target in inflammaging of the pulmonary disease. Prev Med. 2012; 54:S20–28. https://doi.org/10.1016/j.ypmed.2011.11.014 [PubMed]
- 10. Burastero SE, Magnani Z, Confetti C, Abbruzzese L, Oddera S, Balbo P, Rossi GA, Crimi E. Increased expression of the CD80 accessory molecule by alveolar macrophages in asthmatic subjects and its functional involvement in allergen presentation to autologous TH2 lymphocytes. J Allergy Clin Immunol. 1999; 103:1136–42. https://doi.org/10.1016/s0091-6749(99)70189-2 [PubMed]
- 11. Fitzpatrick AM, Holguin F, Teague WG, Brown LA. Alveolar macrophage phagocytosis is impaired in children with poorly controlled asthma. J Allergy Clin Immunol. 2008; 121:1372–78, 1378.e1–3. https://doi.org/10.1016/j.jaci.2008.03.008 [PubMed]
- 12. Wiese AV, Ender F, Quell KM, Antoniou K, Vollbrandt T, König P, Köhl J, Laumonnier Y. The C5a/C5aR1 axis controls the development of experimental allergic asthma independent of LysM-expressing pulmonary immune cells. PLoS One. 2017; 12:e0184956. https://doi.org/10.1371/journal.pone.0184956 [PubMed]
- 13. Staples KJ, Hinks TS, Ward JA, Gunn V, Smith C, Djukanović R. Phenotypic characterization of lung macrophages in asthmatic patients: overexpression of CCL17. J Allergy Clin Immunol. 2012; 130:1404–12.e7. https://doi.org/10.1016/j.jaci.2012.07.023 [PubMed]
- 14. Kitada M, Ogura Y, Koya D. The protective role of Sirt1 in vascular tissue: its relationship to vascular aging and atherosclerosis. Aging (Albany NY). 2016; 8:2290–307. https://doi.org/10.18632/aging.101068 [PubMed]
- 15. Hui X, Zhang M, Gu P, Li K, Gao Y, Wu D, Wang Y, Xu A. Adipocyte SIRT1 controls systemic insulin sensitivity by modulating macrophages in adipose tissue. EMBO Rep. 2017; 18:645–57. https://doi.org/10.15252/embr.201643184 [PubMed]
- 16. Liu L, Zhu X, Zhao T, Yu Y, Xue Y, Zou H. Sirt1 ameliorates monosodium urate crystal-induced inflammation by altering macrophage polarization via the PI3K/Akt/STAT6 pathway. Rheumatology (Oxford). 2019; 58:1674–83. https://doi.org/10.1093/rheumatology/kez165 [PubMed]
- 17. Kim KE, Kim H, Heo RW, Shin HJ, Yi CO, Lee DH, Kim HJ, Kang SS, Cho GJ, Choi WS, Roh GS. Myeloid-specific SIRT1 Deletion Aggravates Hepatic Inflammation and Steatosis in High-fat Diet-fed Mice. Korean J Physiol Pharmacol. 2015; 19:451–60. https://doi.org/10.4196/kjpp.2015.19.5.451 [PubMed]
- 18. Zhang Z, Xu J, Liu Y, Wang T, Pei J, Cheng L, Hao D, Zhao X, Chen HZ, Liu DP. Mouse macrophage specific knockout of SIRT1 influences macrophage polarization and promotes angiotensin II-induced abdominal aortic aneurysm formation. J Genet Genomics. 2018; 45:25–32. https://doi.org/10.1016/j.jgg.2018.01.002 [PubMed]
- 19. Yoshizaki T, Schenk S, Imamura T, Babendure JL, Sonoda N, Bae EJ, Oh DY, Lu M, Milne JC, Westphal C, Bandyopadhyay G, Olefsky JM. SIRT1 inhibits inflammatory pathways in macrophages and modulates insulin sensitivity. Am J Physiol Endocrinol Metab. 2010; 298:E419–28. https://doi.org/10.1152/ajpendo.00417.2009 [PubMed]
- 20. Li Z, Wu Y, Chen HP, Zhu C, Dong L, Wang Y, Liu H, Xu X, Zhou J, Wu Y, Li W, Ying S, Shen H, Chen ZH. MTOR Suppresses Environmental Particle-Induced Inflammatory Response in Macrophages. J Immunol. 2018; 200:2826–34. https://doi.org/10.4049/jimmunol.1701471 [PubMed]
- 21. Qian G, Jiang W, Zou B, Feng J, Cheng X, Gu J, Chu T, Niu C, He R, Chu Y, Lu M. LPS inactivation by a host lipase allows lung epithelial cell sensitization for allergic asthma. J Exp Med. 2018; 215:2397–412. https://doi.org/10.1084/jem.20172225 [PubMed]
- 22. Berger M, de Boer JD, Bresser P, van der Poll T, Lutter R, Sterk PJ, van der Zee JS. Lipopolysaccharide amplifies eosinophilic inflammation after segmental challenge with house dust mite in asthmatics. Allergy. 2015; 70:257–64. https://doi.org/10.1111/all.12544 [PubMed]
- 23. Chen Z, Pan J, Jia Y, Li D, Min Z, Su X, Yuan H, Shen G, Cao S, Zhu L, Wang X. Effect of memory CD4+ T cells’ signal transducer and activator of transcription (STATs) functional shift on cytokine-releasing properties in asthma. Cell Biol Toxicol. 2017; 33:27–39. https://doi.org/10.1007/s10565-016-9357-6 [PubMed]