



Introduction
Despite significant progress in therapeutic strategies has been achieved in recent years, colorectal cancer (CRC) still ranks the third most frequent tumor worldwide, followed by lung and breast carcinoma [1]. The mortality of CRC in developed countries is high, and is keeping an increasing tendency [2]. At present, there are over 1 million newly diagnosed cases each year, seriously threatening People’s health and quality of life [3]. Therefore, in-depth exploration of the process and specific molecular mechanism of CRC is particularly important, which may offer the references or strategies for updated treatments.
Sprouty (SPRY) are initially identified as the inhibitors of receptor tyrosine kinase regulating tracheal branching in Drosophila [4]. SPRY4, a membrane of SPRY family (SPRY1-4), is generally recognized as a regulator of receptor tyrosine kinases [5]. In addition, SPRY4 also exerts regulatory effects on cell growth, differentiation, and metastasis during the development of malignant tumors in various types of cancers [6, 7]. SPRY4 suppresses the migration and stem cell-related properties of breast carcinoma cells and suppresses cell motility of prostate cancer cells [8–10]. Nevertheless, SPRY4 acts as an oncogene to promote ovarian cancer invasion and accelerate human ovarian cancer progression [11]. Thus, SPRY4 plays a dual role in human cancers and the function of SPRY4 often varies among different tumor types. However, up to date, the role of SPRY4 in CRC still remains unclear.
EZH2, a histone-lysine N-methyltransferase enzyme, has been documented to be associated with the occurrence and development of malignant tumors [12–14], and also be involved in the cellular biological functions of various cancers, including gastric cancer, hepatocellular carcinoma, bladder cancer, and nasopharyngeal cancer [15–18]. Furthermore, several studies have reported the abnormally high expression of EZH2, which is also positively correlated with the poor prognosis of CRC [19–21], indicating an important role of EZH2 in CRC.
The aim of the present study is to explore the role of SPRY4 during tumor progression of CRC in vivo and in vitro. In addition, this study also investigates the relationship between SPRY4 and EZH2 to further understand the potential regulatory mechanism.
Results
Differential expression of SPRY4 and EZH2 between colorectal cancer cells and normal human colon cell
To explore the role of SPRY4 in CRC, we first screened the expression level of SPRY4 and EZH2 in four human CRC cell lines, SW620, SW480, LOVO and HCT116, and a normal human colorectal epithelial cell line, NCM460, via western blotting and RT-qPCR. As shown in Figure 1A–1C, the translation and transcription levels of SPRY4 genes between normal cell line and cancer cell lines were statistically different. In particular, NCM460 cell line exhibited the highest level of SPRY4, while SW480 exhibited the lowest. In contrast, the expression of EZH2 gene was upregulated in all cancer cell lines, and SW480 showed the highest level (Figure 1D–1F). Therefore, we selected SW480 cell line to continue the investigation of SPRY4.
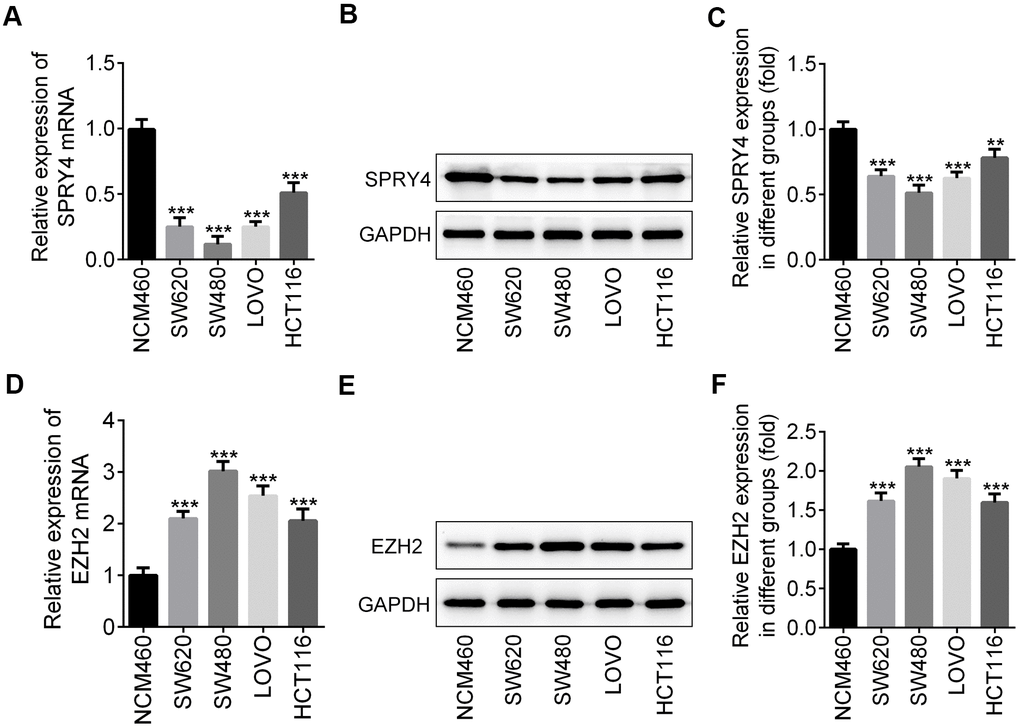
Figure 1. Differential expression of SPRY4 and EZH2 between colorectal cancer cells and normal human colon cell. From four human CRC cell lines, SW620, SW480, LOVO and HCT116, and a normal human colon mucosal epithelial cell line NCM460, we detected the mRNA level and protein expression of SPRY4 using western blotting and RT-qPCR, respectively (A–C). The mRNA level and protein expression of EZH2 were detected using western blotting and RT-qPCR, respectively (D–F). **, ***p<0.01, 0.001 vs NCM460.
Effects of SPRY4 overexpression on CRC cell proliferation
To examine the impacts of SPRY4, we constructed the overexpression plasmid of SPRY4 and the enhanced expression of SPRY4 was observed in transfected SW480 (Figure 2A–2C). Next, the effects of SPRY4 overexpression on SW480 cell proliferation were determined by CCK8, colony formation and EdU assays. As shown in Figure 2D, SPRY4 overexpression dramatically suppressed the proliferation ability of SW480 cells in a time-dependent manner. Additionally, compared with negative control group, the number of cell colonies was also significantly reduced upon SPRY4 overexpression (Figure 2E). Furthermore, EdU assay further revealed that overexpression of SPRY4 obviously decreased the proliferation ability of SW480 cells (Figure 2F).
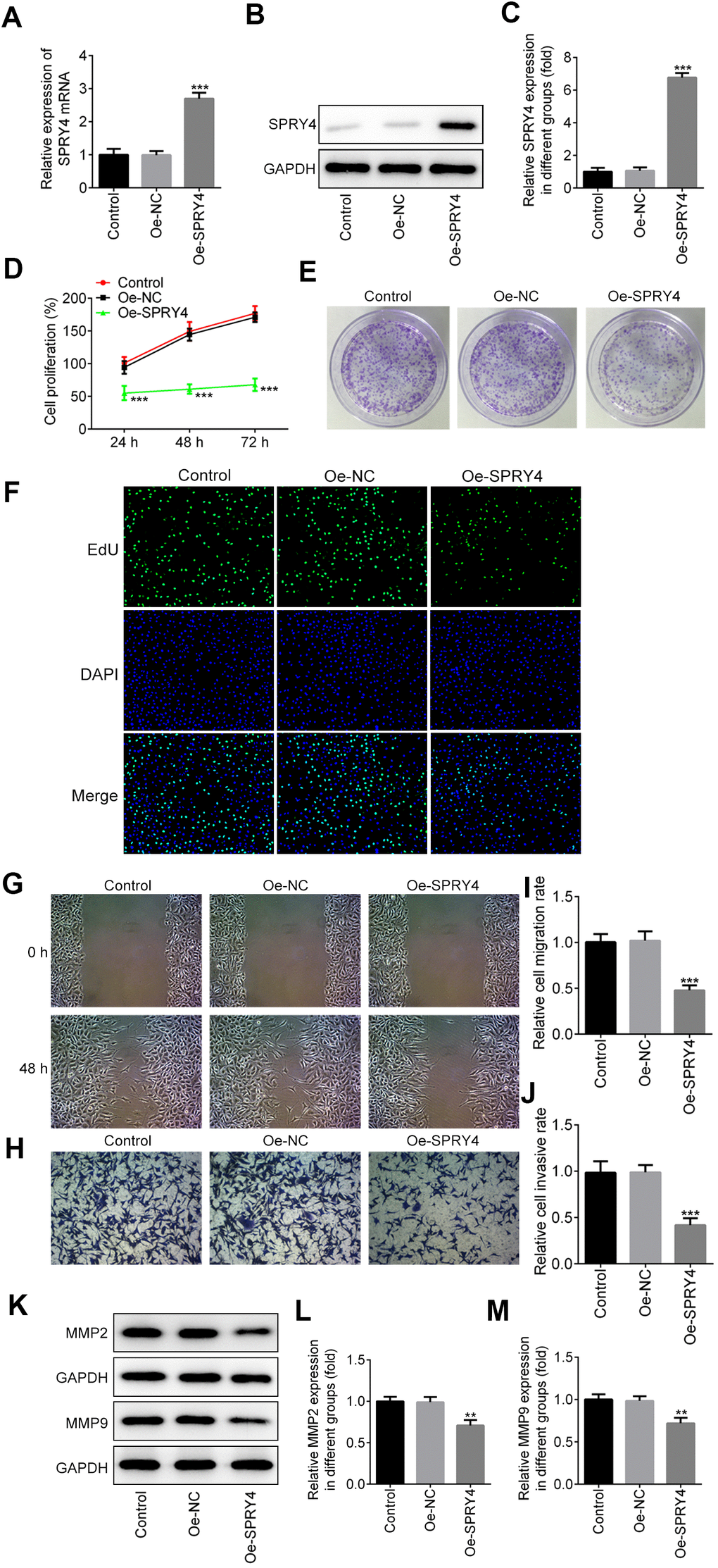
Figure 2. Effects of SPRY4 overexpression on CRC cell proliferation, migration and invasion. The overexpression plasmid of SPRY4 was constructed and transfected into SW480 cells. The mRNA level and protein expression of SPRY4 were detected using RT-qPCR and western blotting, respectively (A–C). Cell proliferation ability was determined using CCK-8 assay (D), cell colony formation (E), and EdU assay (F). Wound-healing and Transwell assays were conducted to measure cell migration and invasion, respectively (G–J). Protein expression of MMP2 and MMP9 was detected by western blotting (K–M). **, ***p<0.01, 0.001 vs Oe-NC.
Effects of SPRY4 overexpression on invasion and migration of CRC cell
We further evaluated the impacts of SPRY4 on CRC cell mobility. As exhibited by the in vitro scratch assay (Figure 2G, 2I), wound closure of SW480 cell was decreased upon SPRY4 overexpression, compared to the control. For the Transwell assay, SW480 cells with different treatments were allowed to invade for 48 h and a similar trend was also observed. The SPRY4 overexpression treatment resulted in the least amount of cancer cells in the lower chamber (Figure 2H, 2J). Since matrix metalloproteinase 2 (MMP2) and MMP9 are the key members of the MMP family which can degrade the extracellular matrix, we continued to evaluate their expression mediated by SPRY4 overexpression with western-blotting assay. As depicted in Figure 2K–2M, the expression of MMP2 and MMP9 was downregulated by SPRY4, which was only about half of that of control group. In general, these data indicated a preventive role of SPRY4 against invasion and migration of CRC cells.
Effect of SPRY4 overexpression on apoptosis in CRC cells
Next, we assessed the impacts of SPRY4 on apoptosis in SW480 cells. As shown in Figure 3A, 3B, compared to the control group, SPRY4 overexpression remarkably improved cell apoptosis rate. In addition, SPRY4 overexpression upregulated the protein expressions of pro-apoptosis-related Bax and cleaved caspase-3 while down-regulated the anti-apoptosis-related Bcl-2 (Figure 3C–3F), which further demonstrated the promotive effect on cell apoptosis in SW480 cells.
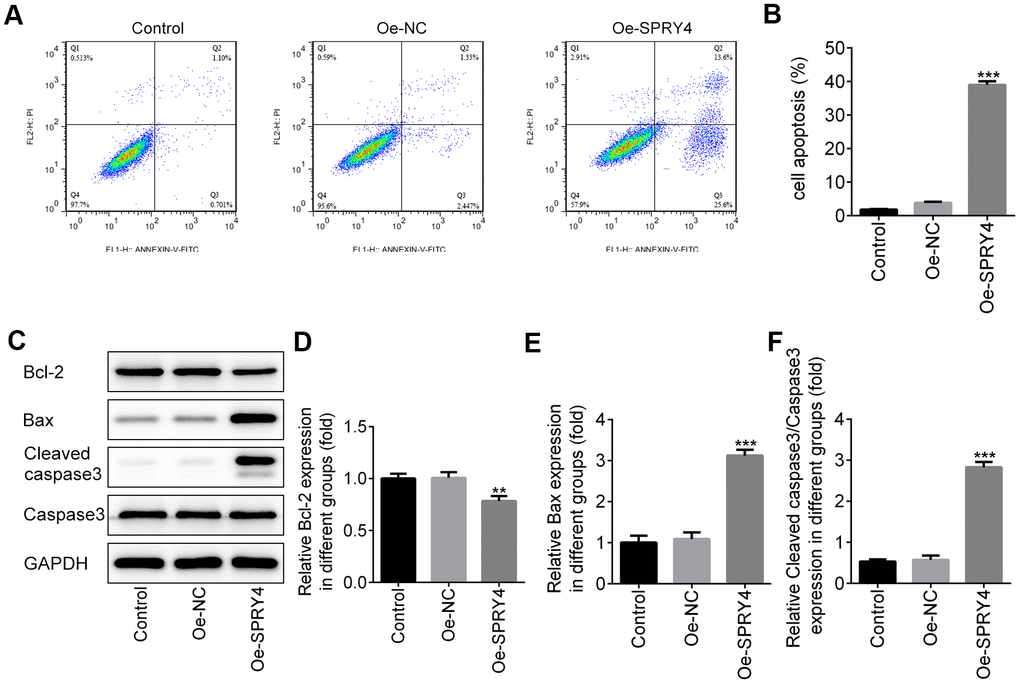
Figure 3. Effects of SPRY4 overexpression on CRC cell apoptosis. The overexpression plasmid of SPRY4 was constructed and transfected into SW480 cells. Then, cell apoptotic rate was analyzed using flow cytometry assay (A, B). Furthermore, the protein expressions of Bcl-2, Bax, Cleaved caspase-3 and caspase-3 were measured using western blotting (C–F). **, ***p<0.01, 0.001 vs Oe-NC.
SPRY4 reduced the expression of EZH2 and MDM2
SW480 cells is a p53 double-mutant cell line, whereas the main target of MDM2 is wild-type p53 [22]. In addition, activated p53 was demonstrated to suppress EZH2 gene expression through repression of the EZH2 gene promoter, and SPRY4 was reported to promote tumor cell proliferation by activating EZH2 [23, 24], thus, the relationship among SPRY4, MDM2 and EZH2 in SW480 cells aroused our interest. Intriguingly, we observed a statistically significant inverse correlation between SPRY4 and the expression of EZH2 and MDM2 (Figure 4A–4C). Upon SPRY4 overexpression, the expression of EZH2 and MDM2 in SW480 were downregulated, which were similar with the treatment of GSK126 at 50 μM, indicating an underlying inhibition role of SPRY4 towards EZH2 and MDM2. Next, the immunoprecipitation assay was used to evaluate the association between EZH2 and MDM2, and western blot was used to measure the expression of EZH2 and MDM2. We found that EZH2 and MDM2 could bind to each other (Figure 4D, 4E). Thus, taking the important role of p53 into consideration, the functional role of SPRY4 in CRC might be achieved partly through EZH2/MDM2/p53 axis.
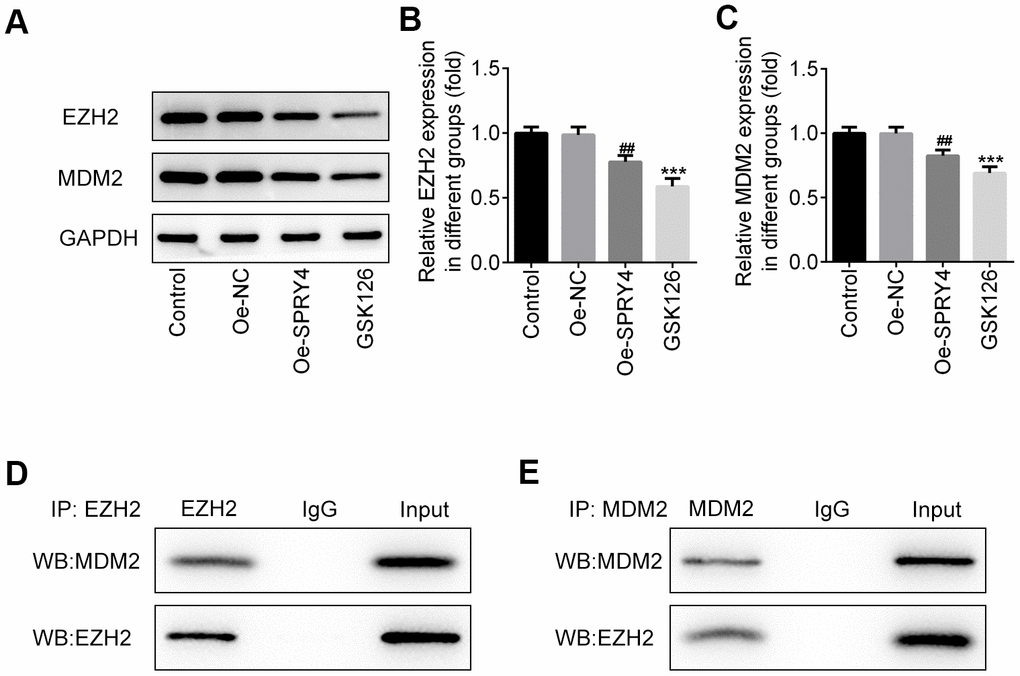
Figure 4. SPRY4 reduced the expression of EZH2 and MDM2. SW480 cells were transfected with overexpression plasmid of SPRY4 or co-treated with overexpression plasmid of SPRY4 and GSK126 (an EZH2 inhibitor). In each group, protein expressions of EZH2 and MDM2 were detected using western blotting (A–C). Immunoprecipitation assay, followed by western blotting, was used to evaluate the association between EZH2 and MDM2 (D, E). ***p<0.001 vs Control; ##p<0.01 vs Oe-NC.
The effects of SPRY4 silencing on proliferation, invasion and apoptosis were partly via regulating EZH2
To further study the impacts of SPRY4, we successfully transfected SW480 cells with si-SPRY4 plasmid (Figure 5A). The protein expression of EZH2 was demonstrated to be up-regulated after transfection with si-SPRY4 (Figure 5B). A series of cellular functional experiments showed that after SPRY4 was knocked down, the proliferation of SW480 cells was greatly enhanced (reached ~200% after 72 h of incubation) and the inhibitory effects of GSK126 on cell proliferation in such cell were compromised. Similar trends were also observed in the wound-healing and Transwell assays. Compared with the GSK126 alone, GSK126 combined with si-SPRY4 transfection enhanced cell migration and invasion (Figure 5D–5G), upregulated expression of MMP2 and MMP9 (Figure 5H–5J), and reduced level of apoptosis (Figure 6A, 6B), as well as upregulated the expression of Bcl-2 and decreased the expression of Bax and cleaved caspase-3 (Figure 6C–6F). These results again highlighted the SPRY4 as a repressor to oncogene EZH2.
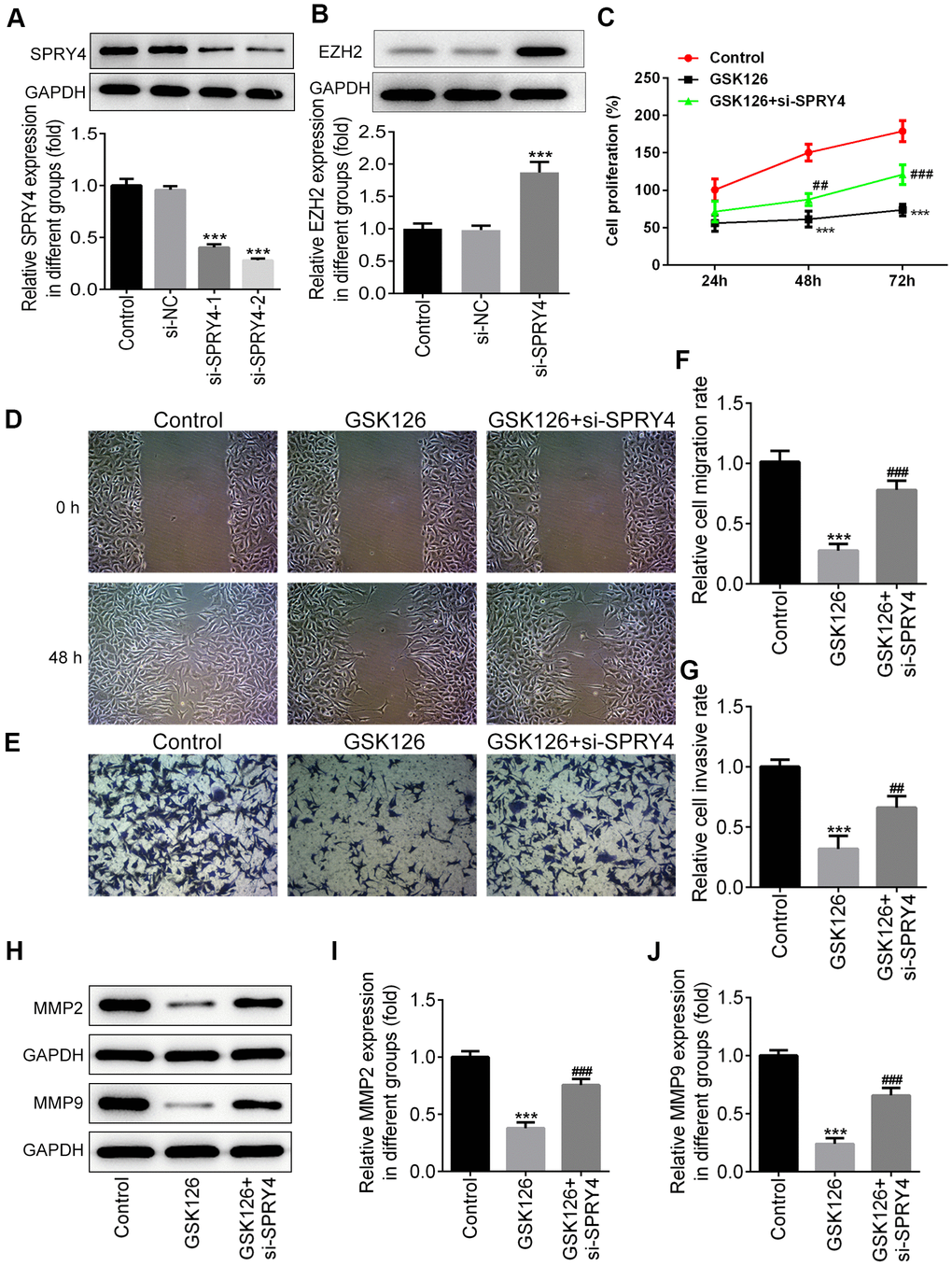
Figure 5. The function of SPRY4 silence on proliferation, migration and invasion in SW480 cells. SW480 cells were transfected with si-SPRY4 plasmid-1/2 and the empty plasmid, then the protein expression of SPRY4 in different groups was measured using western blotting (A). After transfection, the protein expression of EZH2 was also measured using western blot (B). ***P<0.001 vs si-NC. SW480 cells were treated with GSK126 or co-treated with GSK126 and si-SPRY4. CCK-8 assay was conducted to determine cell proliferation ability (C). Wound-healing and Transwell assays were performed to determine cell migration and invasion abilities (D–G). The protein expression of MMP2 and MMP9 was measured using western blotting (H–J). ***p<0.001 vs Control; ##, ###p<0.01, 0.001 vs GSK126.
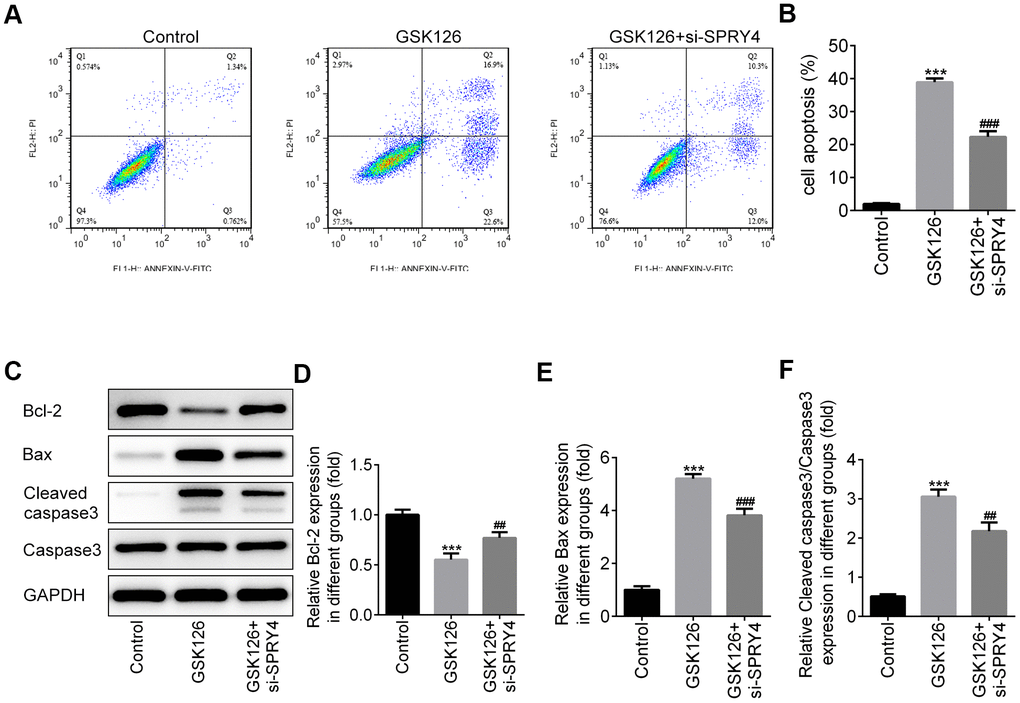
Figure 6. The function of SPRY4 silence on apoptosis in SW480 cells. SW480 cells were treated with GSK126 or co-treated with GSK126 and si-SPRY4. Cell apoptotic rate was analyzed using flow cytometry assay (A, B). Moreover, the protein expressions of Bcl-2, Bax, Cleaved caspase-3 and caspase-3 were measured using western blotting (C–F). ***p<0.001 vs Control; ##, ###p<0.01, 0.001 vs GSK126.
SPRY4 regulated tumorigenesis in vivo
To further investigate the effects of SPRY4 on tumorigenesis in vivo, we have additionally constructed plasmids to overexpress SPRY4 (Oe-SPRY4) or silence SPRY4 (si-SPRY4), respectively. The SW480 cells transfected with Oe-SPRY4 or si-SPRY4 were subcutaneously implanted into the axilla of the nude mice. The weight of mice and the tumor volume were monitored before sacrifice. Our results showed that both tumor size and tumor weight were reduced by Oe-SPRY4 and increased by si-SPRY4, especially, and the promotive effect of si-SPRY4 on tumor size and tumor weight was partly reversed by GSK126 (Figure 7A–7D). The protein expression of EZH2 in tumor tissues was reduced in Oe-SPRY4 group but increased in si-SPRY4 group (Figure 7E, 7F), indicating the involvement of EZH2 in the antitumor activity of SPRY4. Immunohistochemistry assay indicated that the expression of Ki67 was remarkably increased upon si-SPRY4, which was also partly declined upon co-treatment of si-SPRY4 and GSK126 (Figure 7G). Moreover, H&E staining was applied to observe the morphological changes in major organs in different groups. As shown in Figure 7H, the liver, lung and kidney were not injured in each group, indicating that cancer metastasis did not occur. Collectively, our data demonstrated that the SPRY4 regulated tumorigenesis in vivo.
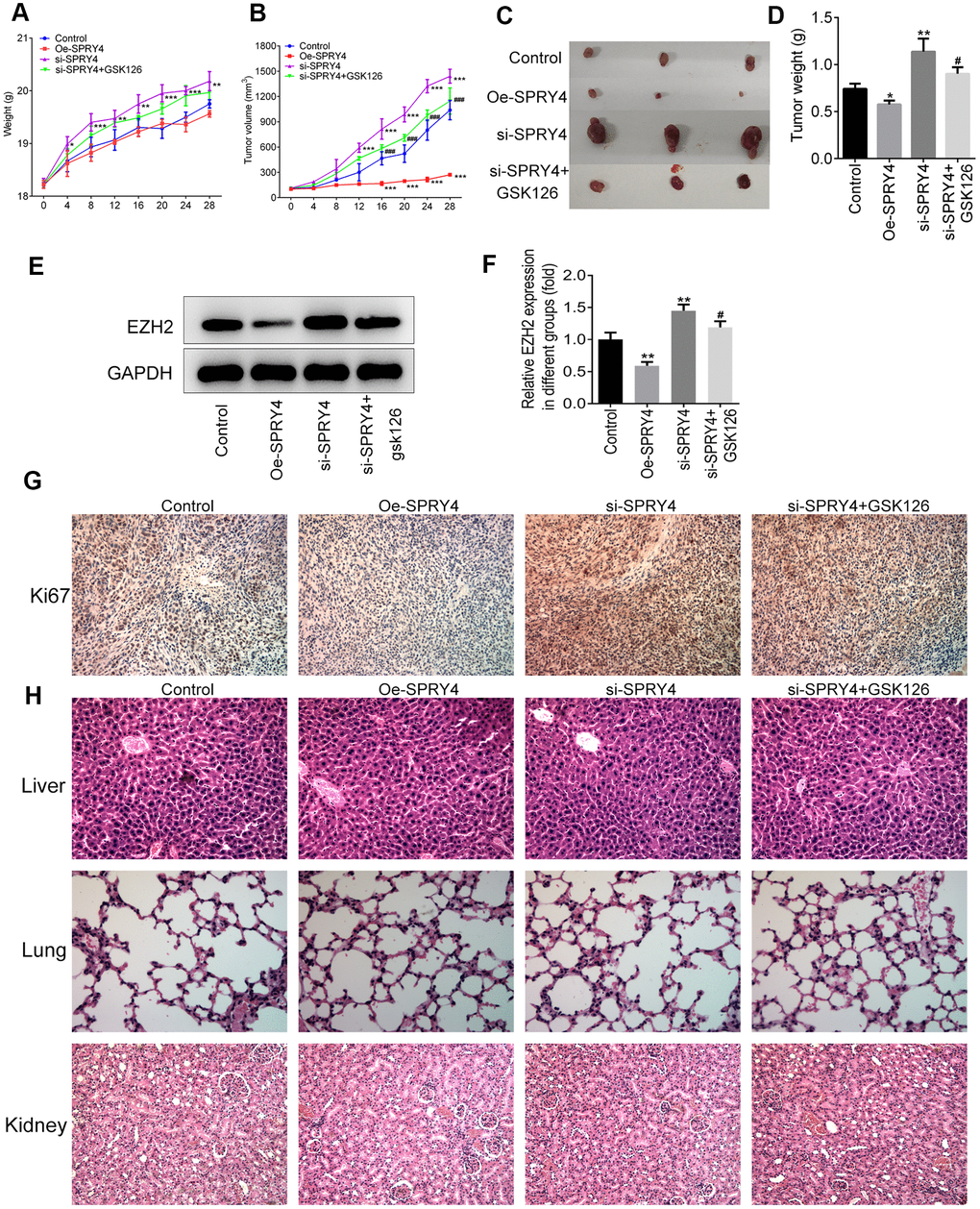
Figure 7. SPRY4 regulated tumorigenesis in vivo. The SW480 cells transfected with Oe-SPRY4 or si-SPRY4 were subcutaneously implanted into the axilla of the nude mice. The weight and tumor volume of mice were monitored every four days (A, B). After sacrifice, the total tumors were separated from the mice and were weighed (C, D). The protein expression of EZH2 in tumor tissues was detected using western blot (E, F). The expression of Ki67 was analyzed by immunohistochemistry (G). This histology of liver, lung and kidney was measured by H&E staining (H). *, **, ***p<0.05, 0.01, 0.001 vs Control; #, ###p<0.05, 0.001 vs si-SPRY4.
Discussion
Pathogenesis and metastasis of CRC generally coincides with the successive dysregulation of certain genes. Even though mounting evidence has highlighted the SPRY4 functions as tumor suppressor, the effects and the mechanism of SPRY4 in regulating CRC progression remain unclear. The purpose of this study is to clarify whether the suppressive role of SPRY4 in tumorigenesis involves modulation of CRC. In order to assess the impacts of SPRY4 on CRC, we first measured the mRNA and protein expression of SPRY4 in CRC cell lines and the normal colorectal cell line. We demonstrated that differential expression modality between cancer cells and normal cells in SPRY4, which hinted that SPRY4 might serve as a tumor suppressor gene that negatively regulates tumor development.
Then, to clarify the role of SPRY4 in CRC, we explored the effects of SPRY4 on CRC cellular biological activities. The uncontrolled proliferation of CRC cells is the main cause for tumor metastasis. Recently, much attention has been paid on the exploration of specific genes or drugs which have inhibitory effects on cell proliferation during the development of CRC. For example, high expression of ubiquitin-like modifier-activating enzyme 2 (UBA2) was reported to be associated with poor prognosis of patients with CRC. Further basic experiments exhibited that downregulated UBA2 could significantly inhibit the proliferation of CRC cell lines in vitro and suppress tumor growth in vivo [25], and butyrate could interrupt the metabolism of CRC cells and ultimately result in the suppression of cell proliferation by targeting pyruvate kinase M2, thus suppressing the development of CRC [26]. These existing evidence revealed that inhibition of cell over-proliferation is an alternative approach to discover the effective treatment for CRC therapy. In the present study, we found that the overexpression of SPRY4 significantly reduced cell proliferation by inhibiting cell viability and hindering colony formation of SW480 cells.
Apoptosis, as a process of the programmed cell death, is essential for the development and for the well-functioning of multicellular systems, and also is an important mechanism among anti-cancer research [27]. In this study, we found that overexpression of SPRY4 significantly promoted cell apoptosis rate of SW480 cells. What’s more, overexpression of SPRY4 showed significant inhibitory effects on the expression of Bcl-2 and promotive effects on the expression of Bax and cleaved caspase-3. Caspase-3 is known as an executioner caspase, and exerts critical effects during cell apoptosis by cleaving many key cellular proteins, no matter through intrinsic mitochondrial pathway or the extrinsic death receptor pathway [28]. Bcl-2 is a critical anti-apoptotic gene which can inhibit cell apoptosis by reducing the generation of reactive oxygen and hindering the release of cytochrome C. While Bax is regarded as a pro-apoptotic gene as the anti-apoptotic activity of Bcl-2 can be abolished by Bax [29]. Thus, these results suggested that the overexpression of SPRY4 might promote apoptosis in SW480 cells by upregulating Bax and caspase-3 while downregulating Bcl-2.
We next investigated the possible targets of SPRY4 in CRC cells through analysis by immunoprecipitation and western blotting. Considering the important role of p53 for SPRY4 and the close relationship among p53, MDM2 and EZH2, it is interesting to find that EZH2 is an appropriate theoretical candidate target of SPRY4. EZH2 is highly expressed in various malignant tumors and is closely involved the occurrence and development of cancers. EZH2 activation can promote expansion of breast tumor initiating cells [30], whereas GSK126, the novel EZH2 inhibitor, exerts anticancer effects on gastric cancer and prostate cancer by inhibiting cell migration, invasion and angiogenesis [31, 32]. In the present study, both the expression of EZH2 and MDM2 were downregulated upon SPRY4 overexpression. Moreover, with the observation that GSK126 could mimic the inhibitory effect of SPRY4 on EZH2 and MDM2, our work indicated that EZH2 might be a direct target of SPRY4 in CRC. EZH2 was verified to bind to MDM2, and MDM2 could bind p53 to promote p53 degradation, which then influenced CRC progression [33, 34]. Thus, the antitumor effects of SPRY4 might be achieved partly by regulating EZH2-mediated MDM2/p53 pathway. The more detailed information concerning on an in-depth research of the mechanism action of SPRY4 is deserved to be investigated in the future.
Collectively, this study found that SPRY4 might act as an anti-tumor gene in CRC and SPRY4 exerted a suppressive effect on CRC progression via the inhibition of EZH2. This study suggests that SPRY4 may serve as a promising candidate of therapeutic targets for CRC treatment.
Materials and Methods
Cells
Human colorectal epithelial cell line NCM460 and colorectal cancer cell lines, including SW620, SW480, LOVO and HCT116, were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA). All cells were cultured in complete Dulbecco’s modified Eagle medium (DMEM; HyClone, Logan, UT) with 10 % fetal bovine serum (FBS; HyClone, Logan, UT).
Quantitative real-time PCR
Trizol reagent (Beyotime, Haimen, China) was applied to extract entire RNA. After determining the quality and concentration of the extracted RNA, 1 μg of RNA was reverse transcribed to complementary DNA (cDNA) by a reverse transcriptase (TaKaRa, Japan). Then, quantitative real time PCR (qRT-PCR) was conducted using the SYBR-Green method (Applied Biosystems, USA) to detect the expression level of corresponding genes following the manufacturer’s guide. The primers used were listed as follows: SPRY4 (forward-5′-TCTGACCAACGGCTCTTAGAC-3′, reverse- 5′-GTGCCATAGTTGACCAGAGTC-3′), EZH2 (forward 5′-TTGTTGGCGGAAGCGTGTAAAATC-3′, reverse-5′-TCCCTAGTCCCGCGCAATGAGC-3′), and β-actin (forward-5′-GCACCACACCTTCTACAATG-3′, reverse-5′-TGCTTGCTGATCCACATCTG). RT-qPCR was performed in triplicate for each sample. The mRNA level of these genes was calculated using 2-ΔΔCT method and normalized to that of β-actin mRNA.
Western blotting
Total protein was extracted from the cells using lysis buffer containing proteinase inhibitor cocktail. The protein concentration was measured using the BCA protein assay kit (Beyotime, Haimen, China). Then, the same amount of protein was resolved by 12% SDS-PAGE and transferred to PVDF membranes, followed by blocking of 5 % skimmed milk for 2 h at room temperature. Subsequently, the membranes were incubated with primary antibodies at 4° C overnight. After horseradish peroxidase (HRP)-conjugated second incubation, the bands were developed by ECL assay (Beyotime, Haimen, China). The antibodies used were as followed: Anti-SPRY4 antibody (ab59785), anti-KMT6/EZH2 antibody (ab186006), anti-MDM2 antibody (ab38618), anti-Bcl-2 antibody (ab182858), anti-Bax antibody (ab32503), anti-MMP2 antibody (ab215986), anti-MMP9 antibody (ab219372), anti-Cleaved Caspase-3 antibody (ab32042), anti-β-actin antibody (ab8227) and Rabbit IgG, monoclonal- Isotype Control (ab172730) were obtained from Abcam (MA, USA).
Cell transfection
Cell transfection was conducted using lipofectamine 2000 (Invitrogen, USA) in accordance with the guide of the manufacturer. The pcDNA3.1-SPRY4 (Oe-SRPY4) and pcDNA3.1 negative control (Oe-NC) vectors were obtained from GenePharma (Shanghai, China). SW480 cells were transfected with 50 nM Oe-SRPY4 and Oe-NC, respectively. 48 after transfection, the transfection efficacy was detected by qRT-PCR and western blotting.
Cell proliferation assay
Cell proliferation was determined using Cell Counting Kit-8 (CCK-8) assay. In brief, the transfected SW480 cells were inoculated into 96-well plates (2000 cells/well) and incubated in a humidified atmosphere with 5% CO2 at 37° C. After incubation for various durations, 10 μl of CCK-8 agent was added into each well for a further 3 h’s incubation. Afterwards, a microplate reader (Bio-Tek, USA) was used to detect the optical density of each well at 450 nm.
Colony formation assay
The transfected SW480 cells were seeded into 6-well plates (400 cells/well) and incubated for 2 weeks. During this period, culture medium was replaced every three days. At last, cells were fixed with 4 % paraformaldehyde and stained with 0.1 % crystal violet to observe the colony formation.
EdU assay
The transfected SW480 cells were inoculated into 24-well plates and cultured in a humidified atmosphere with 5% CO2 at 37° C for 24 h. EdU (50 μmol/L) was added for a further incubation of 8 h. Afterwards, cells were fixed with 4% formaldehyde at room temperature for 15 min, followed by incubation with Triton X-100 for 15 min. Subsequently, cells were incubated with EdU agent and stained with DAPI. Finally, cells were observed under a confocal microscopy (Olympus, Japan).
Wound-healing assay
The transfected SW480 cells were seeded into 6-well plates. When 100% confluence achieved, a scratch was generated by using a 20 μl pipette tip. Then, the culture medium was replaced to DMEM without FBS. Cell images were obtained at 0 and 48 h under a light microscope (Olympus, Japan).
Transwell assay
The transfected SW480 cells suspended in FBS-free DMEM were inoculated into the upper chamber of a 24-well Transwell pre-coated with Matrigel (BD Biosciences, USA) at a density of 4 x 104 cells per well. The complete medium with 10% FBS was added to the lower chamber of the Transwell. 48 h later, the cells on the surface of the upper chamber were wiped out. The invasive cells were fixed with 4 % paraformaldehyde and were stained with 0.1 % crystal violet. The images were visualized using a light microscope (Olympus, Japan).
Flow cytometry analysis
The transfected SW480 cells were collected, washed with cold PBS, and re-suspended in 100 μL of binding buffer. Subsequently, cells were stained with annexin V-FITC and propidium iodide (PI) according to the guide of the manufacturer. The signal was acquired by a FACS caliber flow cytometry and analyzed with FACS Diva software (BD Biosciences, USA).
In vivo tumor xenograft study
Male Balb/c nude mice at the age of 4-6 weeks were obtained from Shanghai SLAC laboratory animal co. Ltd (Shanghai, China). The animal experiments were carried out in accordance with the protocols approved by the Institutional Animal Care and Use Committee of The Affiliated Hospital of Kunming Medical University. SW480 cells were transfected with SPRY4 knockdown plasmid or SPRY4 overexpression plasmid. The transfected cells were subcutaneously injected into the axilla of the mice. GSK126 (an inhibitor of EZH2, Selleck) was administrated intravenously at a dose of 150 mg/kg. The tumor volume was calculated as follows: tumor volume = 1/2 × length ×width2. After 28 days, the mice were euthanized, and the tumors were weighed and collected for further tests.
Hematoxylin-Eosin (H&E) staining
The tumor tissue samples were fixed in 4 % paraformaldehyde, dehydrated in ethanol and embedded in paraffin blocks. Then, the samples were cut into 5-μm-thickness of sections. After staining with H&E reagent, the images were observed under a light microscope for randomly five fields.
Immunohistochemistry
The paraffin-embedded tissue samples were deparaffinized, rehydrated and subjected to antigen retrieval. After washing with PBS and blocked with PBS containing 5% normal goat serum, the sections were washed with PBS and incubated with primary antibody against Ki67 and biotinylated secondary antibody (Santa Cruz, CA, USA). The sections were developed with DAB staining and hematoxylin counterstain. All images were photographed for five randomly fields.
Statistical analysis
Data were presented as mean ± SEM. All the data were analyzed using the SPSS 18.0 statistical software. The differences among groups were determined using one way ANOVA analysis followed by Tukey’s post hoc test. p < 0.05 was considered to indicate a statistically significant difference.
Author Contributions
KY and JG contributed conception and design of the study. JG, HZ, QL, JD and WX performed the experiments and statistical analysis. KY wrote the first draft of the manuscript. JG, HZ and QL wrote sections of the manuscript. All authors contributed to manuscript revision, read and approved the submitted version.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Funding
This study was supported by Yunnan Science and Technology Planning Project (NO. 2017FE467 (-072)).
References
- 1. Siegel R, Ma J, Zou Z, Jemal A. Cancer statistics, 2014. CA Cancer J Clin. 2014; 64:9–29. https://doi.org/10.3322/caac.21208 [PubMed]
- 2. Das V, Kalita J, Pal M. Predictive and prognostic biomarkers in colorectal cancer: A systematic review of recent advances and challenges. Biomed Pharmacother. 2017; 87:8–19. https://doi.org/10.1016/j.biopha.2016.12.064 [PubMed]
- 3. Ferlay J, Soerjomataram I, Dikshit R, Eser S, Mathers C, Rebelo M, Parkin DM, Forman D, Bray F. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer. 2015; 136:E359–86. https://doi.org/10.1002/ijc.29210 [PubMed]
- 4. Hacohen N, Kramer S, Sutherland D, Hiromi Y, Krasnow MA. Sprouty encodes a novel antagonist of FGF signaling that patterns apical branching of the drosophila airways. Cell. 1998; 92:253–63. https://doi.org/10.1016/s0092-8674(00)80919-8 [PubMed]
- 5. Mason JM, Morrison DJ, Basson MA, Licht JD. Sprouty proteins: multifaceted negative-feedback regulators of receptor tyrosine kinase signaling. Trends Cell Biol. 2006; 16:45–54. https://doi.org/10.1016/j.tcb.2005.11.004 [PubMed]
- 6. Masoumi-Moghaddam S, Amini A, Morris DL. The developing story of sprouty and cancer. Cancer Metastasis Rev. 2014; 33:695–720. https://doi.org/10.1007/s10555-014-9497-1 [PubMed]
- 7. Tennis MA, Van Scoyk MM, Freeman SV, Vandervest KM, Nemenoff RA, Winn RA. Sprouty-4 inhibits transformed cell growth, migration and invasion, and epithelial-mesenchymal transition, and is regulated by Wnt7A through PPARgamma in non-small cell lung cancer. Mol Cancer Res. 2010; 8:833–43. https://doi.org/10.1158/1541-7786.MCR-09-0400 [PubMed]
- 8. Vanas V, Mühlbacher E, Kral R, Sutterlüty-Fall H. Sprouty4 interferes with cell proliferation and migration of breast cancer-derived cell lines. Tumour Biol. 2014; 35:4447–56. https://doi.org/10.1007/s13277-013-1587-0 [PubMed]
- 9. Wang J, Thompson B, Ren C, Ittmann M, Kwabi-Addo B. Sprouty4, a suppressor of tumor cell motility, is down regulated by DNA methylation in human prostate cancer. Prostate. 2006; 66:613–24. https://doi.org/10.1002/pros.20353 [PubMed]
- 10. Jing H, Liaw L, Friesel R, Vary C, Hua S, Yang X. Suppression of Spry4 enhances cancer stem cell properties of human MDA-MB-231 breast carcinoma cells. Cancer Cell Int. 2016; 16:19. https://doi.org/10.1186/s12935-016-0292-7 [PubMed]
- 11. So WK, Cheng JC, Liu Y, Xu C, Zhao J, Chang VT, Leung PC. Sprouty4 mediates amphiregulin-induced down-regulation of E-cadherin and cell invasion in human ovarian cancer cells. Tumour Biol. 2016; 37:9197–207. https://doi.org/10.1007/s13277-016-4790-y [PubMed]
- 12. Kuser-Abali G, Gong L, Yan J, Liu Q, Zeng W, Williamson A, Lim CB, Molloy ME, Little JB, Huang L, Yuan ZM. An EZH2-mediated epigenetic mechanism behind p53-dependent tissue sensitivity to DNA damage. Proc Natl Acad Sci USA. 2018; 115:3452–57. https://doi.org/10.1073/pnas.1719532115 [PubMed]
- 13. Zhao Y, Ding L, Wang D, Ye Z, He Y, Ma L, Zhu R, Pan Y, Wu Q, Pang K, Hou X, Weroha SJ, Han C, et al. EZH2 cooperates with gain-of-function p53 mutants to promote cancer growth and metastasis. EMBO J. 2019; 38:e99599. https://doi.org/10.15252/embj.201899599 [PubMed]
- 14. Fillmore CM, Xu C, Desai PT, Berry JM, Rowbotham SP, Lin YJ, Zhang H, Marquez VE, Hammerman PS, Wong KK, Kim CF. EZH2 inhibition sensitizes BRG1 and EGFR mutant lung tumours to TopoII inhibitors. Nature. 2015; 520:239–42. https://doi.org/10.1038/nature14122 [PubMed]
- 15. Chen DL, Ju HQ, Lu YX, Chen LZ, Zeng ZL, Zhang DS, Luo HY, Wang F, Qiu MZ, Wang DS, Xu DZ, Zhou ZW, Pelicano H, et al. Long non-coding RNA XIST regulates gastric cancer progression by acting as a molecular sponge of miR-101 to modulate EZH2 expression. J Exp Clin Cancer Res. 2016; 35:142. https://doi.org/10.1186/s13046-016-0420-1 [PubMed]
- 16. Chen S, Pu J, Bai J, Yin Y, Wu K, Wang J, Shuai X, Gao J, Tao K, Wang G, Li H. EZH2 promotes hepatocellular carcinoma progression through modulating miR-22/galectin-9 axis. J Exp Clin Cancer Res. 2018; 37:3. https://doi.org/10.1186/s13046-017-0670-6 [PubMed]
- 17. Ler LD, Ghosh S, Chai X, Thike AA, Heng HL, Siew EY, Dey S, Koh LK, Lim JQ, Lim WK, Myint SS, Loh JL, Ong P, et al. Loss of tumor suppressor KDM6A amplifies PRC2-regulated transcriptional repression in bladder cancer and can be targeted through inhibition of EZH2. Sci Transl Med. 2017; 9:eaai8312. https://doi.org/10.1126/scitranslmed.aai8312 [PubMed]
- 18. Yu M, Li Y, Li M, Lu D. Eudesmin exerts antitumor effects by down-regulating EZH2 expression in nasopharyngeal carcinoma cells. Chem Biol Interact. 2019; 307:51–57. https://doi.org/10.1016/j.cbi.2019.04.028 [PubMed]
- 19. Huang KB, Zhang SP, Zhu YJ, Guo CH, Yang M, Liu J, Xia LG, Zhang JF. Hotair mediates tumorigenesis through recruiting EZH2 in colorectal cancer. J Cell Biochem. 2019; 120:6071–77. https://doi.org/10.1002/jcb.27893 [PubMed]
- 20. Chen Z, Yang P, Li W, He F, Wei J, Zhang T, Zhong J, Chen H, Cao J. Expression of EZH2 is associated with poor outcome in colorectal cancer. Oncol Lett. 2018; 15:2953–61. https://doi.org/10.3892/ol.2017.7647 [PubMed]
- 21. Wang D, Quiros J, Mahuron K, Pai CC, Ranzani V, Young A, Silveria S, Harwin T, Abnousian A, Pagani M, Rosenblum MD, Van Gool F, Fong L, et al. Targeting EZH2 reprograms intratumoral regulatory T cells to enhance cancer immunity. Cell Rep. 2018; 23:3262–74. https://doi.org/10.1016/j.celrep.2018.05.050 [PubMed]
- 22. Rochette PJ, Bastien N, Lavoie J, Guérin SL, Drouin R. SW480, a p53 double-mutant cell line retains proficiency for some p53 functions. J Mol Biol. 2005; 352:44–57. https://doi.org/10.1016/j.jmb.2005.06.033 [PubMed]
- 23. Zhou M, Zhang XY, Yu X. Overexpression of the long non-coding RNA SPRY4-IT1 promotes tumor cell proliferation and invasion by activating EZH2 in hepatocellular carcinoma. Biomed Pharmacother. 2017; 85:348–54. https://doi.org/10.1016/j.biopha.2016.11.035 [PubMed]
- 24. Tang X, Milyavsky M, Shats I, Erez N, Goldfinger N, Rotter V. Activated p53 suppresses the histone methyltransferase EZH2 gene. Oncogene. 2004; 23:5759–69. https://doi.org/10.1038/sj.onc.1207706 [PubMed]
- 25. He P, Sun X, Cheng HJ, Zou YB, Wang Q, Zhou CL, Liu WQ, Hao YM, Meng XW. UBA2 promotes proliferation of colorectal cancer. Mol Med Rep. 2018; 18:5552–62. https://doi.org/10.3892/mmr.2018.9613 [PubMed]
- 26. Li Q, Cao L, Tian Y, Zhang P, Ding C, Lu W, Jia C, Shao C, Liu W, Wang D, Ye H, Hao H. Butyrate suppresses the proliferation of colorectal cancer cells via targeting pyruvate kinase M2 and metabolic reprogramming. Mol Cell Proteomics. 2018; 17:1531–45. https://doi.org/10.1074/mcp.RA118.000752 [PubMed]
- 27. Ashkenazi A, Fairbrother WJ, Leverson JD, Souers AJ. From basic apoptosis discoveries to advanced selective BCL-2 family inhibitors. Nat Rev Drug Discov. 2017; 16:273–84. https://doi.org/10.1038/nrd.2016.253 [PubMed]
- 28. Riedl SJ, Shi Y. Molecular mechanisms of caspase regulation during apoptosis. Nat Rev Mol Cell Biol. 2004; 5:897–907. https://doi.org/10.1038/nrm1496 [PubMed]
- 29. Reed JC. Proapoptotic multidomain bcl-2/bax-family proteins: mechanisms, physiological roles, and therapeutic opportunities. Cell Death Differ. 2006; 13:1378–86. https://doi.org/10.1038/sj.cdd.4401975 [PubMed]
- 30. Chang CJ, Yang JY, Xia W, Chen CT, Xie X, Chao CH, Woodward WA, Hsu JM, Hortobagyi GN, Hung MC. EZH2 promotes expansion of breast tumor initiating cells through activation of RAF1-β-catenin signaling. Cancer Cell. 2011; 19:86–100. https://doi.org/10.1016/j.ccr.2010.10.035 [PubMed]
- 31. Lin W, Chen Y, Zeng L, Ying R, Zhu F. [Effect of a novel EZH2 inhibitor GSK126 on prostate cancer cells]. Zhejiang Da Xue Xue Bao Yi Xue Ban. 2016; 45:356–63. [PubMed]
- 32. Chen YT, Zhu F, Lin WR, Ying RB, Yang YP, Zeng LH. The novel EZH2 inhibitor, GSK126, suppresses cell migration and angiogenesis via down-regulating VEGF-A. Cancer Chemother Pharmacol. 2016; 77:757–65. https://doi.org/10.1007/s00280-016-2990-1 [PubMed]
- 33. Li H, Zhang J, Tong JH, Chan AW, Yu J, Kang W, To KF. Targeting the oncogenic p53 mutants in colorectal cancer and other solid tumors. Int J Mol Sci. 2019; 20:5999. https://doi.org/10.3390/ijms20235999 [PubMed]
- 34. Deisenroth C, Zhang Y. Ribosome biogenesis surveillance: probing the ribosomal protein-Mdm2-p53 pathway. Oncogene. 2010; 29:4253–60. https://doi.org/10.1038/onc.2010.189 [PubMed]