



Introduction
Inequalities in aging-related health and mortality by aspects of the neighborhood environment are consistently demonstrated in the literature, independent of individual-level socioeconomic position [1–8]. This has significant implications for Black individuals in the United States who are overrepresented in socioeconomically disadvantaged and resource-limited neighborhoods due to historical racialized segregation policies and limited upward residential mobility [9–11]. Despite the robust evidence linking neighborhoods to health and mortality, less is known about the underlying biological mechanisms that may explain how the neighborhood environment becomes physiologically embodied to influence longevity.
Individuals living in disadvantaged neighborhoods are more likely exposed to chronic stressors, including discrimination, crime, and despair, and elevated levels of stress [12, 13]. Chronic elevated stress can detrimentally affect on multiple body systems, including inflammatory, cardiovascular, and neuroendocrine. This cumulative biological wear and tear, or aging, is widely known as allostatic load [14]. Allostatic load is a measure of the biological aging process and several studies have reported associations with adverse neighborhood environments [15, 16]. Emerging evidence also suggests that living in adverse neighborhood environments may be linked to biological aging at the cellular- and molecular-levels [17–24]. Epigenetic mechanisms, specifically DNA methylation (DNAm), offer insights into possible molecular pathways. DNAm regulates gene expression without altering the underlying DNA sequence, is associated with health outcomes and mortality, and responsive to exogenous factors, including area-level environmental exposures [20, 21, 25–27]. Several measures of DNAm aging, also known as “epigenetic clocks,” have been developed using DNA microarray technology and statistical algorithms to identify alterations in DNAm that are highly correlated with chronological age [28–30]. Recently, Levine et al., developed a DNAm aging biomarker to better estimate aging as related to clinical phenotypes of chronic disease [30]. Accelerated DNAm aging, defined as instances where DNAm age is greater than chronological age, has been associated with adverse physical and mental health outcomes and all-cause mortality [31]. Taken together, it is possible that molecular biomarkers, such as DNAm, are useful early indicators of neighborhood quality-related health and aging effects.
Living in areas characterized by exposure to disadvantage [23, 27, 32], violence and crime [33], and air pollution [34] have been shown to accelerate DNAm aging. In studies of neighborhood environment and DNAm aging, neighborhood environment is most often assessed using administrative data sources (i.e. U.S. Census Bureau’s American Community Survey) and boundaries (i.e. census tract). However, an individual’s perception of their neighborhood environment may also capture salient experiences of the neighborhood that impact molecular mechanisms [21]. Positive aspects of the neighborhood, such as perception of neighborhood social cohesion, may act to buffer the effects of an adverse neighborhood environment on DNAm aging, as found in studies relating neighborhoods to health [35–38]. Accordingly, our study sought to investigate both the independent and joint impacts of neighborhood social environment and cohesion on three measures of DNAm aging—Horvath’s epigenetic clock, Hannum’s epigenetic clock, and Levine’s PhenoAge— among a sample of predominately Black adults living in Detroit, MI [28–30]. Given previous work suggesting aging biomarkers differ by sex and with respect to area-level environmental characteristics [17, 34, 39], we investigated associations stratified by sex, as well as in the full sample.
Results
Description of study sample
Chronological age was highly correlated with Horvath’s clock (Pearson r = 0.81), Hannum’s clock (Pearson r = 0.84), and PhenoAge (Pearson r = 0.80; Supplementary Figure 1). Selected baseline characteristics for the 158 study participants included in this analysis are shown in Table 1. Overall, the study sample comprised primarily of Black (87%) and women (61%) participants. Participants were long-term residents of their neighborhoods (mean = 18.2 ± 16.5 years). The mean proportion of residents living below the federal poverty level was 38.1% ± 9.3%, while the mean social cohesion score was 8.1 (SD = 2.1). On average, men were 4.5 years younger than women (men: 50.2 ± 14.5 years; women: 54.7 ± 12.8 years). For both men and women, DNAm age was higher for Horvath’s clock (men: 58.6 ± 11.2 years; women: 62.1 ± 11.2 years) and Hannum’s clock (men: 56.8 ± 12.2; women: 59.8 ± 10.6 years) and lower for PhenoAge clock (men: 47.2 ± 13.9; women: 53.3 ± 13.2 years) than chronological age.
Table 1. Selected sociodemographic and lifestyle characteristics of the 158 participants included in the study (Detroit Neighborhood Health Study; DNHS).
| Overall (n=158) | Women (n=96) | Men (n=62) | |
| Chronological age in years, mean ± SD | 54.0 ± 13.6 | 54.7 ± 12.8 | 50.2 ± 14.5 |
| Race, N (%) | |||
| Black | 138 (87) | 82 (85) | 56 (90) |
| Non-Black | 20 (12.7) | 14 (14.6) | 6 (9.7) |
| Education, N (%) | |||
| ≤High school | 81 (51) | 40 (42) | 41 (66) |
| >High school | 77 (48.7) | 56 (58.3) | 21 (33.9) |
| Employment status, N (%) | |||
| Unemployed | 109 (69.4) | 65 (67.7) | 44 (72.1) |
| Employed | 48 (31) | 31 (32) | 17 (27) |
| Lifetime smoking status | |||
| Never smoker | 40 (25.3) | 28 (29.2) | 12 (19.4) |
| Ever smoker | 118 (75) | 68 (71) | 50 (81) |
| Lifetime alcohol intake, N (%) | |||
| Never drinker | 31 (19.6) | 23 (24.0) | 8 (12.9) |
| Ever drinker | 127 (80) | 73 (76) | 54 (87) |
| Years lived in current neighborhood, mean ± SD | 18.2 ± 16.5 | 18.2 ± 16.4 | 18.2 ± 16.9 |
| Neighborhood poverty, mean ± SD | 38.1 ± 9.3 | 37.8 ± 9.9 | 37.1 ± 8.6 |
| Neighborhood social cohesion score, mean ± SD | 8.1 ± 2.1 | 8.2 ± 2.2 | 8.1 ± 2.1 |
| Horvath DNA methylation age, mean ± SD | 60.7 ± 11.5 | 62.1 ± 11.2 | 58.6 ± 11.2 |
| Hannum DNA methylation age, mean ± SD | 58.6 ± 11.3 | 59.8 ± 10.6 | 56.8 ± 12.2 |
| Levine’s PhenoAge, mean ± SD | 50.9 ± 13.8 | 53.3 ± 13.2 | 47.2 ± 13.9 |
Neighborhood poverty and DNAm age
We observed evidence to suggest neighborhood poverty accelerated DNAm aging that was primarily driven by the association among women (Figure 1A). After adjusting for factors included in the full model, the pattern of association among women signaled that living in neighborhoods with higher poverty was associated with PhenoAge acceleration (β = 1.4; 95% CI: -0.4, 3.3), which was not seen among men (β = -0.3; 95% CI: -2.2, 1.5). However, the confidence intervals for both women and men included the null. (Supplementary Table 3).
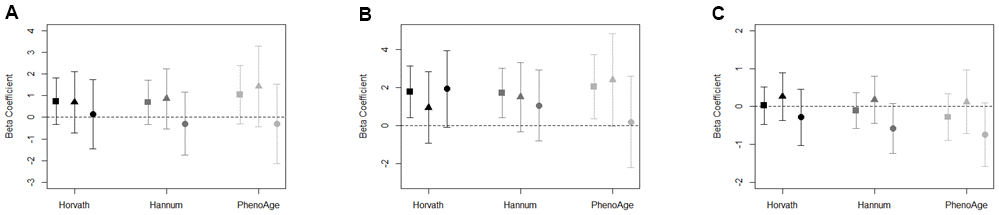
Figure 1. (A–C) Association between (A) neighborhood poverty, (B) neighborhood PC7, (C) neighborhood social cohesion, and DNAm age acceleration measures for total sample (square), women (triangle), and men (circle). Models adjusted for race/ethnicity, education level, employment, smoking status, alcohol intake, and years residing in current neighborhood. Black symbols represent associations with Horvath age acceleration, dark gray represent Hannum age acceleration, and light gray represent PhenoAge acceleration.
Objective neighborhood observations (PCs) and DNAm age
To reduce the number of tests conducted on our limited sample size, we summarized the 19 observed indicators of neighborhood quality using principal components analysis (PCA). Of the eight principal components (PCs) retained, only PC7 was associated with accelerated DNAm aging across each of the three measures of epigenetic aging (Supplementary Table 4). Among the total sample, PC7 was associated with accelerated DNAm aging for Horvath age acceleration (β = 1.8; 95% CI: 0.4, 3.1), Hannum age acceleration (β = 1.7; 95% CI: 0.4, 3.0), and PhenoAge acceleration (β = 2.1; 95% CI: 0.4, 3.8) (Figure 1B). The top positive loadings for PC7 were factors characterized by the presence of abandoned cars and people on the street (Supplementary Table 2). Among women, we observed PhenoAge acceleration in response to living in a neighborhood characterized by PC7 (β = 2.4; 95% CI: -0.0, 4.9), and, among men, suggestion of Horvath age acceleration (β = 1.9; 95% CI: -0.1, 4.0; Supplementary Table 4).
Neighborhood social cohesion and DNAm age
The associations in the full sample between neighborhood social cohesion and DNAm aging biomarkers were largely null (Figure 1C). However, potential differences by sex were observed. For instance, among men, the effect estimate for neighborhood social cohesion and Hannum age acceleration (β = -0.6; 95% CI: -1.2, 0.1) and PhenoAge acceleration (β = -0.7; 95% CI: -1.6, 0.1) were negative; whereas, the effect estimates for Hannum age acceleration and PhenoAge acceleration among women were relatively null (Supplementary Table 3). For both Hannum age and PhenoAge acceleration, the effect estimates for men were outside of the confidence intervals for women, which suggests potential differences by sex.
To assess whether objective neighborhood quality-DNAm aging associations differed by perception of neighborhood social cohesion in the total sample, we conducted stratified analyses in the full models comparing associations among residents living in neighborhoods with higher vs. lower social cohesion scores (based on the median value of 8.1). Our results were suggestive of a difference in the neighborhood PC7-DNAm age acceleration association by neighborhood social cohesion (Figure 2). We found that associations between PC7 and DNAm age acceleration remained elevated for participants living in neighborhoods with lower social cohesion (Horvath: β = 2.1; 95%. CI: 0.6, 3.6; Hannum: β = 2.2; 95%. CI: 0.6, 3.8; PhenoAge: β = 2.3; 95%. CI: 0.3, 4.7); however, there appeared to be no association among participants living in neighborhoods with higher neighborhood social cohesion (Figure 2 and Supplementary Table 5). Similarly, for neighborhood poverty and DNAm age acceleration, we observed evidence of associations among participants living in neighborhoods with lower social cohesion, while null associations were observed among residents living in neighborhoods with higher social cohesion (Supplementary Figure 2).
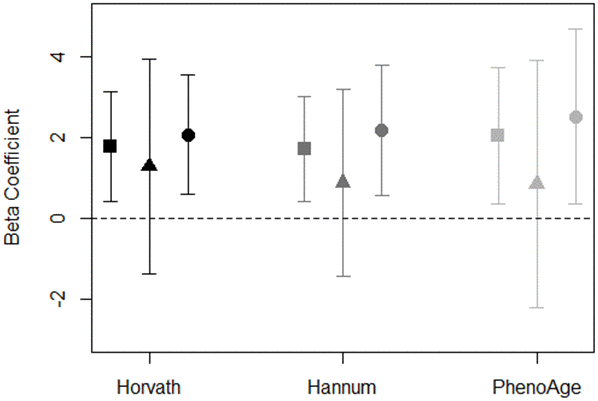
Figure 2. Association of PC7 and DNAm age acceleration measures stratified by neighborhood social cohesion for total sample (square), high social cohesion (triangle), and low social cohesion (circle). Models adjusted for race/ethnicity, education level, employment, smoking status, alcohol intake, and years residing in current neighborhood. Black symbols represent associations with Horvath age acceleration, dark gray represent Hannum age acceleration, and light gray represent PhenoAge acceleration.
Because inflammation is one mechanism through which the neighborhood environment may influence DNAm aging, we further examined the impact of blood immune cell counts on our associations. For each exposure-outcome association, adjusting for cell type proportions attenuated our associations, suggesting that inflammation may partially explain associations between neighborhood attributes and DNAm aging (Supplementary Tables 3–5).
Discussion
In this study of predominately Black adults living in Detroit, we observed evidence of potential links between aspects of the neighborhood social environment and accelerated DNAm aging that were primarily driven by associations in women. PhenoAge outpaced chronological age by nearly 2 years for women living in neighborhoods characterized by increased presence of abandoned cars and people on the street, as well as with higher levels of poverty. On the other hand, neighborhood social cohesion appeared to be associated with negative DNAm age acceleration (or DNAm age deceleration) for men, but not women. When examined together in models stratified on perceptions of neighborhood social cohesion, associations between measures of neighborhood environment (poverty and PC7) and DNAm age acceleration remained elevated for participants living in neighborhoods with lower social cohesion, while associations were relatively null for those residing in neighborhoods with high social cohesion. Taken together, it is possible that despite living in neighborhoods with higher poverty and lower quality, social cohesion may act to protect against accelerating DNAm aging.
Our findings are consistent with previous research linking neighborhood disadvantage to accelerated DNAm aging in children and adults [27, 32]. In our study, we found that women’s DNAm aging was more sensitive to neighborhood poverty and PC7 than men. Similarly, a prior study among 100 African American women found that a standard deviation increase in neighborhood disadvantage was associated with a 9-month increase (β = 0.75; 95% CI: 0.27, 1.23) in Hannum’s age acceleration [27], which is comparable to the 12-month increase in Hannum’s age acceleration we observed in relation to a standard deviation increase in neighborhood poverty among women in our study. Additionally, we explored the impact of neighborhood poverty on a more recent measure of epigenetic aging, PhenoAge acceleration, which estimates aging based on epigenetic loci associated with 10 age-related clinical measures [30]. Neighborhood poverty appears to have a greater effect on PhenoAge acceleration than Hannum’s age acceleration in our study. Many of the clinical measures included in the PhenoAge estimator, such as glucose levels [40] and C-reactive protein [41–43], have demonstrated associations with neighborhood-level social characteristics. This might possibly explain the stronger associations observed for PhenoAge than the other epigenetic aging biomarkers in this and previous studies of the neighborhood social environment.
Unlike previous studies, our study used direct observations of neighborhood characteristics conducted by trained evaluators, which provide an objective assessment of the neighborhood’s natural social environment [44]. We found that PC7, defined by the presence of people on the street and abandoned cars, was associated with accelerated DNAm aging across each of the three measures of epigenetic aging examined in this study. This finding is consistent with our previous work showing neighborhood quality measures, namely abandoned cars, people on the street, and non-art graffiti, were associated with an epigenetic biomarker of mortality [23].
Neighborhood social cohesion has also been shown to impact physical and mental health outcomes and may have effects at the molecular-level [21, 35–38]. In a study of 1,226 adults in the Multi-Ethnic Study of Atherosclerosis (MESA), the neighborhood social environment, measured as a summary score that included social cohesion, was associated with DNAm in stress- and inflammation-related genes [21]. In our study, men living in neighborhoods with higher neighborhood social cohesion appeared to experience negative Hannum and Pheno age acceleration. Furthermore, in our full models stratified by higher vs. lower neighborhood social cohesion, the impact of PC7 and poverty on DNAm aging remained detrimental among participants living in neighborhoods with lower social cohesion, but not among those living in neighborhoods with higher social cohesion. Additional studies in larger prospective cohorts are needed to corroborate our findings; yet, if replicated our results have important implications for mortality and longevity. Evidence exists demonstrating that neighborhood social cohesion is associated with lower risk of mortality [1]. Given the link between accelerated DNAm aging and mortality, it is possible residing in a neighborhood with greater social cohesion may serve as a buffer to the negative effects of neighborhood deprivation on mortality through decelerated DNAm aging as demonstrated in our study.
Our study examines the impact of neighborhood-level social processes on individual-level molecular mechanisms. Several potential mechanisms exist that may explain molecular response to the neighborhood environment. First, participants residing in neighborhoods with high levels of disadvantage and disorder may experience chronic activation and dysregulation of stress-response and inflammatory pathways and altered DNAm patterns, as suggested in previous studies demonstrating associations between neighborhood disadvantage and DNAm of genes involved in stress-response and inflammation [20, 21]. We observed attenuated results in our models when blood immune cell proportions were included further implicating inflammatory pathways as a mechanistic link. Additional pathways, such as smoking status and dietary behaviors, may affect these associations. Unfortunately, we did not have information on dietary habits, but did adjust for smoking and alcohol consumption in our models, which did not impact our results. There may be additional factors correlated with neighborhood disadvantage like environmental air quality that may explain our associations and should be explored in future investigations. We also observed suggestive differences in patterns of associations for men and women. While these findings warrant follow-up in larger samples, the null associations observed between neighborhood poverty and PC7 with DNAm aging acceleration among men may be explained by the overall health of men in our small study sample. For instance, PhenoAge (a marker of the aging process for key clinical measures of chronic disease) was lower among men in our study than women, which may be a signal that men were in better overall health, and DNAm aging was not responsive to the adverse effects of living in neighborhoods with poor economic and social conditions. Future studies should examine this further.
Our study is not without limitations. Due to our relatively small sample size these analyses should be considered somewhat exploratory in nature. We did not impose a multiple testing correction as the sample size was limited and the outcomes were correlated, as well as biologically-related. In addition, due to the limited sample size, we were unable to perform formal tests of significant differences or explore potentially salient higher-order interactions, such as three-way interactions among sex, neighborhood disadvantage, and social cohesion, which are likely relevant, but cannot be addressed using small sample sizes. Despite our sample size, we were able to pick up signals of associations in the entire cohort and when stratifying on sex and neighborhood social cohesion based on descriptive interpretations, such as confidence interval non-overlap. We focused on summary measures of objective neighborhood quality as these summary measures combine correlated features of the underlying inputs, which can increase power to detect associations. The Detroit Neighborhood Health Study (DNHS) is a population-based study of individuals residing in the Detroit metropolitan area, which may limit the generalizability of our study results to other parts of the United States. However, our findings are in line with previous studies of neighborhood disadvantage and DNAm aging. Detroit was one of the harder hit areas during the U.S. Great Recession from 2007 to 2009, which coincided with the DNHS. While participants in the neighborhoods were long-term residents, we do not have repeated measures of neighborhood quality and are unable to examine the effects of neighborhood changes on DNAm. Lastly, aging is a life course process. The biological aging process is likely a response to the accumulation of environmental insults over one’s life span rather than short-term exposures. Because our analysis was cross-sectional with neighborhood measures and DNAm assessed at similar points in time, we were unable to make inferences on the temporal mechanisms. However, it is unlikely that individuals were selected into their neighborhoods based on their underlying epigenetic age, particularly given that most individuals resided in their homes for over a decade. When combined with the average age of individuals (54.0 years), this would indicate that most individuals began residing in their homes at an age prior to when age (or epigenetic age) driven functional deficits would appear, which would have been the most likely causal mechanism for epigenetic age acceleration to drive neighborhood choice.
In summary, our study explored multiple measures of neighborhood environment, including U.S. Census data for neighborhood poverty, direct observations of neighborhood quality, and survey responses of neighborhood social cohesion, in relation to DNAm aging. We found that individuals living in neighborhoods characterized by higher levels of poverty and the presence of abandoned cars and people on the street experience accelerated aging, where their DNAm age is higher than their chronological age. However, effects of neighborhood poverty and quality on DNAm aging may be buffered by increased neighborhood social cohesion. Given that racial/ethnic minorities are more likely to reside in deprived and disadvantaged neighborhoods, our findings offer a molecular insight into potential mechanisms of health disparities. We suggest that future studies fruitfully interrogate these associations to build upon our understanding of the biosocial mechanisms that contribute to racial/ethnic disparities in health.
Materials and Methods
Study population
Baseline data from adults participating in the DNHS were used for the present study. The DNHS is a population-based prospective cohort study of primarily Black adult residents (18 years of age or older) living in Detroit, MI. Study participants completed annual structured telephone interviews from 2008 through 2012 to assess perceptions of neighborhoods, mental and physical health status, social support, and alcohol and tobacco use. Biospecimens were also collected annually through 2013. A total of 2,081 adults participated in the DNHS, of which 612 agreed to provide either a venipuncture or blood spot at baseline. DNA methylation was measured from whole blood in 179 participants as part of a pilot study, of which 158 with both DNA methylation and neighborhood quality data at baseline examination were included this analysis. Compared to the total DNHS population (n=2,081), participants in our study sample (n=158) were slightly older (52.9 years vs. 50.5 years, p=0.04) and a higher proportion were unemployed (69.4% vs. 58%, p<0.01), but differences in length of residence in the neighborhood were not found (18.2 years vs. 18.2 years, p=0.98). Informed consent was received from all participants in the study prior to participation and the DNHS was approved by the institutional review boards at the University of Michigan (HUM00014138) and the University of North Carolina at Chapel Hill (13-3999).
Neighborhood environment assessment
Neighborhood environment was assessed using information from the U.S. American Community Survey (ACS) participant responses, objective neighborhood evaluations, and participant’s questionnaire responses. To estimate neighborhood poverty, we used data from the 2008-2013 ACS. Neighborhood poverty was defined by the percent of individuals below the federal poverty level. Neighborhood estimates at the census block-group level were aggregated to derive measures within the 54 historically defined neighborhoods in Detroit [45]. Neighborhood poverty was converted to z-scores for this analysis.
Objective neighborhood measures, conducted by trained personnel, were captured from structured assessments of Detroit’s 54 neighborhoods. Within the 54 neighborhoods, 135 block groups were assessed between June and July 2008. During this time, 19 neighborhood characteristics (Supplementary Table 1) were evaluated using a standardized instrument adapted from the New York Social Environment Study to be relevant to Detroit [46]. Evaluators responded with “yes” or “no” and the frequency of “yes” responses were calculated for each variable by block group, which were then averaged by neighborhood. Because our sample size is limited and we know neighborhood characteristics are correlated, we conducted a principal components (PCs) analysis to minimize the number of statistical tests performed. As previously described in a recent study of neighborhood quality and epigenetic mortality risk score, we made the a priori decision to retain the first eight PC that explained 90% of the variance (Supplementary Table 2) [23].
During the first telephone interview, participants were asked to report perceptions of their neighborhood community. Participants were asked about the following characteristics: 1) a close-knit or unified neighborhood; 2) people around here are willing to help their neighbors; 3) people in this neighborhood generally don’t get along with each other (reverse scored); 4) people in this neighborhood do not share the same values (reverse scored); 5) people in this neighborhood can be trusted. Each response was coded from strongly disagree (0) to strongly agree (3). If a participant responded neither disagree nor agree, it was coded as missing. This only impacted three of the 158 participant responses in the total sample. All responses were summed to create a score of individual perception of neighborhood social cohesion, which ranged from 0 to 15 with higher scores indicating greater perception of neighborhood social cohesion. To generate a measure of neighborhood-level perception of social cohesion, individual scores were aggregated and mean neighborhood social cohesion was assigned for each participant living within the same neighborhood [47, 48].
DNAm age variables
Peripheral blood DNA was extracted using venipuncture and genome-wide DNA methylation was measured in whole blood leukocytes using the Illumina Infinium HumanMethylation 450k array using published methods [49]. Samples were bisulfite converted using the EZ-96 DNA methylation kit (Zymo Research). Sample quality control protocol excluded samples with probe detection call rates < 90% and those with an average intensity value of either < 50% or sample mean < 2,000 arbitrary units (AU). Quality control was performed using the R package CpGassoc [50]. Probes with detection p-values > 0.001 and samples with missing data for > 10% of probes were also removed, along with known SNPs and cross-hybridizing probes [51]. Probe normalization was performed using the beta-mixture quantile normalization method using the R package wateRmelon [52, 53]. Following normalization, ComBat was used to account for batch effects using M values converted from the beta values [54]. M values were converted back to beta values for calculation of DNAm age variables.
Horvath’s epigenetic clock, Hannum’s epigenetic clock, and Levine’s PhenoAge were calculated using their published algorithms [28–30]. Horvath’s epigenetic clock consists of 353 CpG sites and is applicable across multiple sources of cells, tissues, and organs across the entire lifespan, including whole blood in adults [29]. Hannum’s epigenetic clock is a single tissue estimator derived from 71 CpG sites from DNA of adult whole blood samples [28]. Horvath’s and Hannum’s clocks are both highly correlated with chronological age; however, weaker associations are observed with clinical characteristics of physiological dysfunction [31]. Levine’s PhenoAge measure was developed using 513 epigenetic loci associated with 10 clinical phenotypes, – albumin, creatinine, glucose, C-reactive protein, lymphocyte percent, red blood cell volume, red cell distribution, alkaline phosphatase, white blood cell count, and chronological age – which were further validated for associations with mortality, co-morbid disease burden, and physical function [30]. The 513 epigenetic loci were converted into an epigenetic clock using regression modeling as done for previous clocks. Levine’s PhenoAge estimator (and PhenoAge acceleration) strongly predicts several aging-related outcomes including all-cause mortality, cancer, coronary heart disease, and Alzheimer’s disease. Each measure of DNAm age was strongly correlated with chronological age in our study population (Supplementary Figure 1A–1C). For the purpose of this analysis, DNAm age residuals were calculated by regressing each DNAm age variable on chronological age resulting in positive and negative deviations from chronological years of age. Positive scores (residuals from the regression model) reflected accelerated DNAm aging.
Additional covariates
Baseline demographic, social, and behavioral factors associated with neighborhood quality and DNAm aging were collected, including chronological age, race/ethnicity, educational attainment, employment status, and number of years lived in current neighborhood. Chronological age was self-reported and included in models as a continuous variable. Race/ethnicity, educational attainment, and employment status were also self-reported and were categorized as shown in Table 1. Participants reported the number of years lived in current neighborhood, which was treated as a continuous variable. Current smoking status (never, ever, current) and lifetime alcohol intake (ever vs. never) were also included in the models.
Statistical analysis
Distributions of selected baseline covariates were examined. Three sex-specific age acceleration variables – Horvath age acceleration (from Horvath’s clock), Hannum age acceleration (from Hannum’s clock), and PhenoAge acceleration (from Levine’s PhenoAge measure) – were derived from regression residuals for the full sample and separately for men and women. We examined whether each neighborhood environment measure – neighborhood poverty, objective neighborhood evaluation (PCs), and neighborhood social cohesion — were independently associated with accelerated DNAm aging. Two separate models were used to estimate our associations of interest: (1) the unadjusted model of neighborhood quality and DNAm age acceleration; and (2) the full model adjusted for race/ethnicity, educational attainment, employment status, lifetime smoking, lifetime alcohol intake, and number of years residing in current neighborhood. We also examined these associations stratified by sex. In a second set of analyses, we assessed whether perception of neighborhood social cohesion provided a buffering effect of the neighborhood poverty and quality measures on DNAm aging. To estimate this, full sample models were stratified by a binary indicator of neighborhood social cohesion, categorized as higher vs. lower social cohesion using the median value. We did not examine this association in the sex-specific models due to our limited sample sizes. Also, because of our limited sample size, nesting within neighborhoods was low (median: 3, IQR: 2-5) with eight neighborhoods only having one participant; therefore, clustering of DNA methylation outcomes by neighborhood is unlikely. Therefore, we modeled our neighborhood variables as individual-level exposures. Given the small sample size and correlations, as well as biological relations, among the epigenetic aging outcome measures we did not impose a multiple testing correction. Blood immune cell counts can serve as markers of inflammation to act as mediators partially explaining associations between neighborhood quality and DNAm aging. To account for this, we estimated blood immune cell proportions using the Houseman method [55] and assessed their influence on our associations in sensitivity analyses. All analyses were performed in R version 3.6 [56]. Results are reported using regression coefficient (β) and associated 95% confidence interval (CI).
Author Contributions
CLM conceived the project idea, developed the study design, conducted the analysis, and drafted the manuscript. All other authors reviewed and provided feedback on the manuscript and approved the final draft for submission.
Acknowledgments and Funding
We are grateful for the time and effort of the DNHS study participants, team, and volunteers. The study authors would also like to acknowledge Sandra Alvarez and Cierra Dungee for their preliminary work on epigenetic aging and neighborhood principal components. The DNHS project was supported by multiple grants from the National Institutes of Health (R01D022720, R01DA022720-S1, RC1MH088283, R01MD011728). CLM received salary and training support from Eunice Kennedy Shriver National Institute of Child Health and Human Development grant T32HD007168, K99MD012808, and R01MD013349. TMZ received support from an NSF Career Award (grant no. 1845796). This manuscript does not necessarily represent the views and policies of the US Environmental Protection Agency. Any mention of tradenames or copyright does not constitute endorsement by the US Environmental Protection Agency.
Conflicts of Interest
Dr. Ward-Caviness is an advisor to the Clock Foundation. The Clock Foundation had no role in this study including the study design, data analysis, and interpretation, or stated conclusion. Other co-authors have no conflicts of interest to report.
References
- 1. Meijer M, Röhl J, Bloomfield K, Grittner U. Do neighborhoods affect individual mortality? A systematic review and meta-analysis of multilevel studies. Soc Sci Med. 2012; 74:1204–12. https://doi.org/10.1016/j.socscimed.2011.11.034 [PubMed]
- 2. Barber S, Hickson DA, Kawachi I, Subramanian SV, Earls F. Neighborhood disadvantage and cumulative biological risk among a socioeconomically diverse sample of African American adults: an examination in the Jackson heart study. J Racial Ethn Health Disparities. 2016; 3:444–56. https://doi.org/10.1007/s40615-015-0157-0 [PubMed]
- 3. Barber S, Hickson DA, Wang X, Sims M, Nelson C, Diez-Roux AV. Neighborhood disadvantage, poor social conditions, and cardiovascular disease incidence among African American adults in the Jackson heart study. Am J Public Health. 2016; 106:2219–26. https://doi.org/10.2105/AJPH.2016.303471 [PubMed]
- 4. Diez Roux AV, Merkin SS, Arnett D, Chambless L, Massing M, Nieto FJ, Sorlie P, Szklo M, Tyroler HA, Watson RL. Neighborhood of residence and incidence of coronary heart disease. N Engl J Med. 2001; 345:99–106. https://doi.org/10.1056/NEJM200107123450205 [PubMed]
- 5. Kawachi I, Berkman LF. Neighborhoods and Health. Oxford University Press; 2003. 369 p. https://doi.org/10.1093/acprof:oso/9780195138382.001.0001
- 6. Li X, Sundquist K, Sundquist J. Neighborhood deprivation and prostate cancer mortality: a multilevel analysis from Sweden. Prostate Cancer Prostatic Dis. 2012; 15:128–34. https://doi.org/10.1038/pcan.2011.46 [PubMed]
- 7. Major JM, Doubeni CA, Freedman ND, Park Y, Lian M, Hollenbeck AR, Schatzkin A, Graubard BI, Sinha R. Neighborhood socioeconomic deprivation and mortality: NIH-AARP diet and health study. PLoS One. 2010; 5:e15538. https://doi.org/10.1371/journal.pone.0015538 [PubMed]
- 8. Nelson K, Schwartz G, Hernandez S, Simonetti J, Curtis I, Fihn SD. The association between neighborhood environment and mortality: results from a national study of veterans. J Gen Intern Med. 2017; 32:416–22. https://doi.org/10.1007/s11606-016-3905-x [PubMed]
- 9. Logan JR. The persistence of segregation in the 21st century metropolis. City Community. 2013; 12:10.1111. https://doi.org/10.1111/cico.12021 [PubMed]
- 10. Sharkey P. The Intergenerational Transmission of Context. American Journal of Sociology. 2008; 113:931–69. https://doi.org/10.1086/522804
- 11. Williams DR, Collins C. Racial residential segregation: a fundamental cause of racial disparities in health. Public Health Rep. 2001; 116:404–16. https://doi.org/10.1093/phr/116.5.404 [PubMed]
- 12. Ross CE, Mirowsky J. Neighborhood disadvantage, disorder, and health. J Health Soc Behav. 2001; 42:258–76. [PubMed]
- 13. Sampson RJ, Raudenbush SW, Earls F. Neighborhoods and violent crime: a multilevel study of collective efficacy. Science. 1997; 277:918–24. https://doi.org/10.1126/science.277.5328.918 [PubMed]
- 14. McEwen BS. Protective and damaging effects of stress mediators. N Engl J Med. 1998; 338:171–79. https://doi.org/10.1056/NEJM199801153380307 [PubMed]
- 15. Ribeiro AI, Amaro J, Lisi C, Fraga S. Neighborhood socioeconomic deprivation and allostatic load: A scoping review. Int J Environ Res Public Health. 2018; 15:1092. https://doi.org/10.3390/ijerph15061092 [PubMed]
- 16. Ribeiro AI, Fraga S, Kelly-Irving M, Delpierre C, Stringhini S, Kivimaki M, Joost S, Guessous I, Gandini M, Vineis P, Barros H. Neighbourhood socioeconomic deprivation and allostatic load: a multi-cohort study. Sci Rep. 2019; 9:8790. https://doi.org/10.1038/s41598-019-45432-4 [PubMed]
- 17. Gebreab SY, Riestra P, Gaye A, Khan RJ, Xu R, Davis AR, Quarells RC, Davis SK, Gibbons GH. Perceived neighborhood problems are associated with shorter telomere length in African American women. Psychoneuroendocrinology. 2016; 69:90–97. https://doi.org/10.1016/j [PubMed]
- 18. Needham BL, Carroll JE, Diez Roux AV, Fitzpatrick AL, Moore K, Seeman TE. Neighborhood characteristics and leukocyte telomere length: the Multi-Ethnic Study of Atherosclerosis. Health Place. 2014; 28:167–72. https://doi.org/10.1016/j.healthplace.2014.04.009 [PubMed]
- 19. Park M, Verhoeven JE, Cuijpers P, Reynolds CF III, Penninx BW. Where You Live May Make You Old: The Association between Perceived Poor Neighborhood Quality and Leukocyte Telomere Length. PLoS One. 2015; 10:e0128460. https://doi.org/10.1371/journal.pone.0128460 [PubMed]
- 20. Reuben A, Sugden K, Arseneault L, Corcoran DL, Danese A, Fisher HL, Moffitt TE, Newbury JB, Odgers C, Prinz J, Rasmussen LJ, Williams B, Mill J, Caspi A. Association of neighborhood disadvantage in childhood with DNA methylation in young adulthood. JAMA Netw Open. 2020; 3:e206095. https://doi.org/10.1001/jamanetworkopen.2020.6095 [PubMed]
- 21. Smith JA, Zhao W, Wang X, Ratliff SM, Mukherjee B, Kardia SLR, Liu Y, Roux AVD, Needham BL. Neighborhood characteristics influence DNA methylation of genes involved in stress response and inflammation: The Multi-Ethnic Study of Atherosclerosis. Epigenetics. 2017; 12:662–73. https://doi.org/10.1080/15592294.2017.1341026 [PubMed]
- 22. Theall KP, Brett ZH, Shirtcliff EA, Dunn EC, Drury SS. Neighborhood disorder and telomeres: connecting children's exposure to community level stress and cellular response. Soc Sci Med. 2013; 85:50–58. https://doi.org/10.1016/j.socscimed.2013.02.030 [PubMed]
- 23. Ward-Caviness CK, Pu S, Martin CL, Galea S, Uddin M, Wildman DE, Koenen K, Aiello AE. Epigenetic predictors of all-cause mortality are associated with objective measures of neighborhood disadvantage in an urban population. Clin Epigenetics. 2020; 12:44. https://doi.org/10.1186/s13148-020-00830-8 [PubMed]
- 24. King KE, Kane JB, Scarbrough P, Hoyo C, Murphy SK. Neighborhood and Family Environment of Expectant Mothers May Influence Prenatal Programming of Adult Cancer Risk: Discussion and an Illustrative DNA Methylation Example. Biodemography Soc Biol. 2016; 62:87–104. https://doi.org/10.1080/19485565.2015.1126501 [PubMed]
- 25. Gondalia R, Baldassari A, Holliday KM, Justice AE, Méndez-Giráldez R, Stewart JD, Liao D, Yanosky JD, Brennan KJ, Engel SM, Jordahl KM, Kennedy E, Ward-Caviness CK, et al. Methylome-wide association study provides evidence of particulate matter air pollution-associated DNA methylation. Environ Int. 2019; 132:104723. https://doi.org/10.1016/j.envint.2019.03.071 [PubMed]
- 26. Gruzieva O, Xu CJ, Breton CV, Annesi-Maesano I, Antó JM, Auffray C, Ballereau S, Bellander T, Bousquet J, Bustamante M, Charles MA, de Kluizenaar Y, den Dekker HT, Duijts L, et al. Epigenome-Wide Meta-Analysis of Methylation in Children Related to Prenatal NO2 Air Pollution Exposure. Environ Health Perspect. 2017; 125:104–10. https://doi.org/10.1289/EHP36 [PubMed]
- 27. Lei MK, Simons RL, Beach SR, Philibert RA. Neighborhood disadvantage and biological aging: using marginal structural models to assess the link between neighborhood census variables and epigenetic aging. J Gerontol B Psychol Sci Soc Sci. 2019; 74:e50–59. https://doi.org/10.1093/geronb/gbx015 [PubMed]
- 28. Hannum G, Guinney J, Zhao L, Zhang L, Hughes G, Sadda S, Klotzle B, Bibikova M, Fan JB, Gao Y, Deconde R, Chen M, Rajapakse I, et al. Genome-wide methylation profiles reveal quantitative views of human aging rates. Mol Cell. 2013; 49:359–67. https://doi.org/10.1016/j.molcel.2012.10.016 [PubMed]
- 29. Horvath S. DNA methylation age of human tissues and cell types. Genome Biol. 2013; 14:R115. https://doi.org/10.1186/gb-2013-14-10-r115 [PubMed]
- 30. Levine ME, Lu AT, Quach A, Chen BH, Assimes TL, Bandinelli S, Hou L, Baccarelli AA, Stewart JD, Li Y, Whitsel EA, Wilson JG, Reiner AP, et al. An epigenetic biomarker of aging for lifespan and healthspan. Aging (Albany NY). 2018; 10:573–91. https://doi.org/10.18632/aging.101414 [PubMed]
- 31. Horvath S, Raj K. DNA methylation-based biomarkers and the epigenetic clock theory of ageing. Nat Rev Genet. 2018; 19:371–84. https://doi.org/10.1038/s41576-018-0004-3 [PubMed]
- 32. Marini S, Davis KA, Soare TW, Zhu Y, Suderman MJ, Simpkin AJ, Smith ADA, Wolf EJ, Relton CL, Dunn EC. Adversity exposure during sensitive periods predicts accelerated epigenetic aging in children. Psychoneuroendocrinology. 2020; 113:104484. https://doi.org/10.1016/j.psyneuen.2019.104484 [PubMed]
- 33. Jovanovic T, Vance LA, Cross D, Knight AK, Kilaru V, Michopoulos V, Klengel T, Smith AK. Exposure to Violence Accelerates Epigenetic Aging in Children. Sci Rep. 2017; 7:8962. https://doi.org/10.1038/s41598-017-09235-9 [PubMed]
- 34. Ward-Caviness CK, Nwanaji-Enwerem JC, Wolf K, Wahl S, Colicino E, Trevisi L, Kloog I, Just AC, Vokonas P, Cyrys J, Gieger C, Schwartz J, Baccarelli AA, et al. Long-term exposure to air pollution is associated with biological aging. Oncotarget. 2016; 7:74510–25. https://doi.org/10.18632/oncotarget.12903 [PubMed]
- 35. Barber S, Hickson DA, Kawachi I, Subramanian SV, Earls F. Double-jeopardy: The joint impact of neighborhood disadvantage and low social cohesion on cumulative risk of disease among African American men and women in the Jackson Heart Study. Soc Sci Med. 2016; 153:107–15. https://doi.org/10.1016/j.socscimed.2016.02.001 [PubMed]
- 36. Dawson CT, Wu W, Fennie KP, Ibañez G, Cano MÁ, Pettit JW, Trepka MJ. Perceived neighborhood social cohesion moderates the relationship between neighborhood structural disadvantage and adolescent depressive symptoms. Health Place. 2019; 56:88–98. https://doi.org/10.1016/j.healthplace.2019.01.001 [PubMed]
- 37. Fone D, White J, Farewell D, Kelly M, John G, Lloyd K, Williams G, Dunstan F. Effect of neighbourhood deprivation and social cohesion on mental health inequality: a multilevel population-based longitudinal study. Psychol Med. 2014; 44:2449–600. https://doi.org/10.1017/S0033291713003255 [PubMed]
- 38. Fone D, Dunstan F, Lloyd K, Williams G, Watkins J, Palmer S. Does social cohesion modify the association between area income deprivation and mental health? a multilevel analysis. Int J Epidemiol. 2007; 36:338–45. https://doi.org/10.1093/ije/dym004 [PubMed]
- 39. McCrory C, Fiorito G, McLoughlin S, Polidoro S, Cheallaigh CN, Bourke N, Karisola P, Alenius H, Vineis P, Layte R, Kenny RA. Epigenetic clocks and allostatic load reveal potential sex-specific drivers of biological aging. J Gerontol A Biol Sci Med Sci. 2020; 75:495–503. https://doi.org/10.1093/gerona/glz241 [PubMed]
- 40. Clark CR, Ommerborn MJ, Hickson DA, Grooms KN, Sims M, Taylor HA, Albert MA. Neighborhood disadvantage, neighborhood safety and cardiometabolic risk factors in African Americans: biosocial associations in the Jackson heart study. PLoS One. 2013; 8:e63254. https://doi.org/10.1371/journal.pone.0063254 [PubMed]
- 41. Assibey-Mensah V, Fabio A, Mendez DD, Lee PC, Roberts JM, Catov JM. Neighbourhood assets and early pregnancy cardiometabolic risk factors. Paediatr Perinat Epidemiol. 2019; 33:79–87. https://doi.org/10.1111/ppe.12531 [PubMed]
- 42. Broyles ST, Staiano AE, Drazba KT, Gupta AK, Sothern M, Katzmarzyk PT. Elevated c-reactive protein in children from risky neighborhoods: evidence for a stress pathway linking neighborhoods and inflammation in children. PLoS One. 2012; 7:e45419. https://doi.org/10.1371/journal.pone.0045419 [PubMed]
- 43. Keita AD, Judd SE, Howard VJ, Carson AP, Ard JD, Fernandez JR. Associations of neighborhood area level deprivation with the metabolic syndrome and inflammation among middle- and older- age adults. BMC Public Health. 2014; 14:1319. https://doi.org/10.1186/1471-2458-14-1319 [PubMed]
- 44. Sampson RJ, Raudenbush SW. Systematic Social Observation of Public Spaces: A New Look at Disorder in Urban Neighborhoods. American Journal of Sociology. 1999; 105:603–51. https://doi.org/10.1086/210356
- 45. Hill EJ, Gallagher J. AIA Detroit: The American Institute of Architects Guide to Detroit Architecture. Detroit, MI: Wayne State University Press. 2002.
- 46. Lê F, Ahern J, Galea S. Neighborhood education inequality and drinking behavior. Drug Alcohol Depend. 2010; 112:18–26. https://doi.org/10.1016/j.drugalcdep.2010.05.005 [PubMed]
- 47. Echeverría S, Diez-Roux AV, Shea S, Borrell LN, Jackson S. Associations of neighborhood problems and neighborhood social cohesion with mental health and health behaviors: the Multi-Ethnic Study of Atherosclerosis. Health Place. 2008; 14:853–65. https://doi.org/10.1016/j.healthplace.2008.01.004 [PubMed]
- 48. Johns LE, Aiello AE, Cheng C, Galea S, Koenen KC, Uddin M. Neighborhood social cohesion and posttraumatic stress disorder in a community-based sample: findings from the detroit neighborhood health study. Soc Psychiatry Psychiatr Epidemiol. 2012; 47:1899–906. https://doi.org/10.1007/s00127-012-0506-9 [PubMed]
- 49. Koenen KC, Uddin M, Chang SC, Aiello AE, Wildman DE, Goldmann E, Galea S. SLC6A4 methylation modifies the effect of the number of traumatic events on risk for posttraumatic stress disorder. Depress Anxiety. 2011; 28:639–47. https://doi.org/10.1002/da.20825 [PubMed]
- 50. Barfield RT, Kilaru V, Smith AK, Conneely KN. CpGassoc: an R function for analysis of DNA methylation microarray data. Bioinformatics. 2012; 28:1280–81. https://doi.org/10.1093/bioinformatics/bts124 [PubMed]
- 51. Chen YA, Lemire M, Choufani S, Butcher DT, Grafodatskaya D, Zanke BW, Gallinger S, Hudson TJ, Weksberg R. Discovery of cross-reactive probes and polymorphic CpGs in the illumina infinium HumanMethylation450 microarray. Epigenetics. 2013; 8:203–09. https://doi.org/10.4161/epi.23470 [PubMed]
- 52. Pidsley R, Y Wong CC, Volta M, Lunnon K, Mill J, Schalkwyk LC. A data-driven approach to preprocessing illumina 450K methylation array data. BMC Genomics. 2013; 14:293. https://doi.org/10.1186/1471-2164-14-293 [PubMed]
- 53. Teschendorff AE, Marabita F, Lechner M, Bartlett T, Tegner J, Gomez-Cabrero D, Beck S. A beta-mixture quantile normalization method for correcting probe design bias in Illumina Infinium 450 k DNA methylation data. Bioinformatics. 2013; 29:189–96. https://doi.org/10.1093/bioinformatics/bts680 [PubMed]
- 54. Leek JT, Johnson WE, Parker HS, Jaffe AE, Storey JD. The sva package for removing batch effects and other unwanted variation in high-throughput experiments. Bioinformatics. 2012; 28:882–83. https://doi.org/10.1093/bioinformatics/bts034 [PubMed]
- 55. Houseman EA, Accomando WP, Koestler DC, Christensen BC, Marsit CJ, Nelson HH, Wiencke JK, Kelsey KT. DNA methylation arrays as surrogate measures of cell mixture distribution. BMC Bioinformatics. 2012; 13:86. https://doi.org/10.1186/1471-2105-13-86 [PubMed]
- 56. R Core Development Team. R: A language and environment for statistical computing. Vienna, Austria. 2019.