



Introduction
Obstructive sleep apnea (OSA), a common chronic sleep disorder, is known to affect millions of people worldwide [1]. Intermittent hypoxia (IH) is the hallmark feature of OSA, which is likely to be the underlying cause for OSA-related comorbidities [2, 3]. Meanwhile, it is noteworthy that OSA is an independent risk for cardiovascular diseases, such as systemic arterial hypertension [4], atrial fibrillation [5], ischemic heart disease [6], metabolic disorders [4], and congestive heart failure [7]. Therefore, the understanding of the molecular mechanisms of OSA-related cardiovascular diseases and development of novel therapeutic strategies for such patients are urgently required.
microRNAs (miRNAs), a cluster of highly conserved non-coding RNAs, play a ubiquitous and key role in the regulation of gene transcription. miRNAs can modulate gene expression through binding to the 3’-UTR region of the target genes, thus resulting in either translational inhibition or degradation of the target genes [8, 9]. It has been reported that miRNAs are implicated in a wide range of cellular processes, including cell viability, development, differentiation, proliferation, and apoptosis [10, 11]. Accumulating evidence has also indicated that miRNAs played crucial roles in multiple heart diseases, such as cardiac hypertrophy [12], heart failure [13], and myocardial infarction [14]. In the past few years, miRNA expression profiles in OSA-related heart disease have been gradually revealed [15, 16]. However, the mechanism underlying the ameliorative effect of miRNAs on IH-induced H9c2 cardiomyocyte injury has not been fully elucidated.
In the current study, we established a model of IH in H9c2 cells. We found that IH could induce cardiomyocyte injury in vitro and lead to miR-3574 downregulation. miR-3574 upregulation ameliorated IH-induced cardiomyocyte injury through regulating cell viability and apoptosis via Axin1, indicating that miR-3574 and Axin1 can serve as a potential novel and effective therapeutic strategy for OSA-related cardiovascular diseases.
Results
IH induces cell injury in H9c2 cardiomyocytes
Following IH treatment, the cell viability of H9c2 cardiomyocytes was then measured by use of CCK-8 assay. The results demonstrated that the level of cell viability was remarkably suppressed following IH treatment in the H9c2 cells (Figure 1A). Furthermore, the expression levels of certain apoptosis-associated proteins, such as Bax, Bcl-2 and caspase-3, were also detected. As shown in Figure 1B, 1C, the protein expression level of Bcl-2 was significantly downregulated, while the expression of Bax and caspase-3 were markedly upregulated following IH treatment in the H9c2 cardiomyocytes.
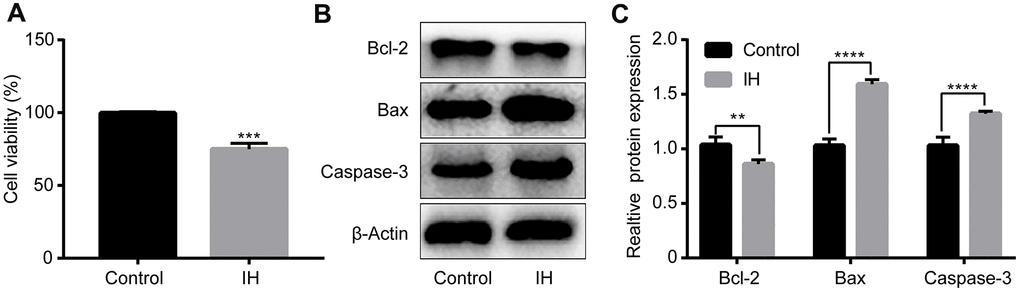
Figure 1. Intermittent hypoxia induces H9c2 cell injury. H9c2 cells were cultured under IH. (A) Cell viability were evaluated by CCK8 assay. (B, C) Cell apoptosis-related proteins were tested by western blotting assays. IH: Intermittent hypoxia. Data are represented as the mean ± SD from three independent experiments. * p < 0.05, ** p < 0.01.
IH reduces miR-3574 expression in H9c2 cardiomyocytes
miR-3574 expression in IH treated H9c2 cells was explored by RT-qPCR. The results revealed that miR-3574 expression was significantly suppressed following IH treatment of H9c2 cells (Figure 1A), indicating that IH treatment reduced the expression of miR-3574 in H9c2 cardiomyocytes. To further determine the role of miR-3574, miR-3574 was overexpressed or inhibited by transfecting with miR-3574 mimic or miR-3574 inhibitor. As shown in Figure 2B, miR-3574 mimic transfection markedly increased miR-3574 expression in the H9c2 cardiomyocytes, while miR-3574 inhibitor transfection significantly decreased miR-3574 expression in the H9c2 cardiomyocytes. These results indicated that transfection efficiency was achieved.
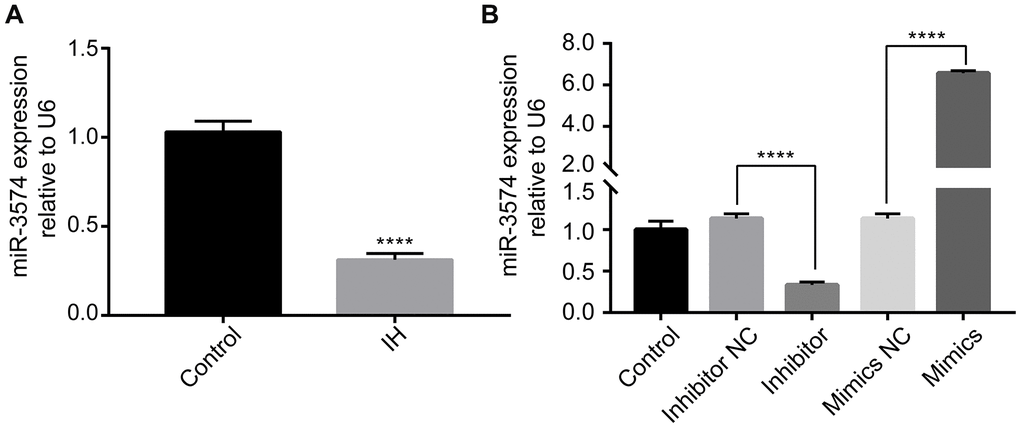
Figure 2. IH reduces miR-3574 expression, and miR-3574 is differentially expressed in H9c2 cardiomyocytes after cell transfection. (A) The expression of miR-3574 was detected by RT-qPCR. (B) Cells were transfected with miR-3574 inhibitor, miR-3574 mimics, and scrambled control. Relative miR-3574 expression was normalized to U6. IH: intermittent hypoxia. Data are represented as the mean ± SD from three independent experiments. * p < 0.05, ** p < 0.01.
Upregulation of miR-3574 attenuates IH-induced cells injury in H9c2 cardiomyocytes
To investigate whether miR-3574 overexpression could protect the cardiomyocytes from IH-induced cells damage, H9C2 cardiomyocytes were transfected by miR-3574 mimics before IH treatment. As shown in Figure 3A, the cell viability decreased in IH-induced cardiomyocytes was increased by miR-3574 upregulation. To further determine the anti-apoptotic effects of miR-3574, the expression levels of apoptosis-associated proteins, including Bax, Bcl-2, and Caspase-3, were also examined. As we expected, western blot assay found that miR-3574 overexpression could significantly decrease the expression of Caspase-3 and Bax and increase the expression of Bcl-2 in IH-injured H9c2 cells. Altogether, these results demonstrate that miR-3574 has an anti-apoptotic role in IH-induced cardiomyocytes injury.
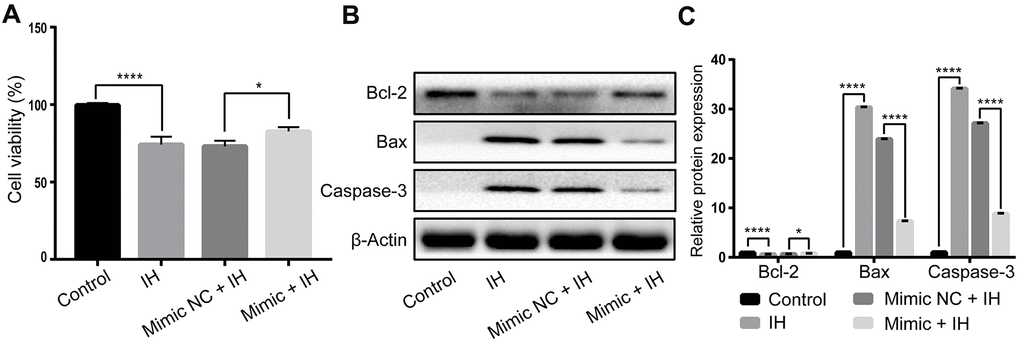
Figure 3. miR-3574 attenuates IH-induced cells injury in H9c2 cardiomyocytes. H9c2 cells were exposed to IH after transfection of miR-3574 mimics, and negative control. (A) Cell viability. (B, C) The expression of apoptosis-related proteins. NC: negative control. IH: intermittent hypoxia. Data are represented as the mean ± SD from three independent experiments. * p < 0.05, ** p < 0.01.
miR-3574 directly targeted Axin1, and inhibited Axin1 expression
Next, we sought to verify the putative targets of miR-3574 in H9c2 cells by IH treatment. We searched for the potential target genes of miR-3574 by the use of PicTar, miRanda, and TargetScan. Bioinformatics analysis identified that Axin1 may be one of the target genes (Figure 4A). To validate it, a dual-luciferase reporter assay was performed. As shown in Figure 4B, miR-3574 significantly reduced the luciferase activity of wild-type Axin1-3’-UTR, but had no effect on that of the mutant one. We then examined the regulation of miR-3574 on Axin1 expression in H9c2 cells. Western blot analysis and RT-qPCR were conducted to test the expression level of Axin1. Our results indicated that both mRNA (Figure 4C) and protein (Figure 4D, 4E) levels of Axin1 were apparently increased in H9c2 cells under IH exposure, which was remarkably suppressed by miR-3574 mimics. Therefore, our findings demonstrate that miR-3574 can directly target the 3’-UTR of Axin1 to inhibit its expression in H9c2 cardiomyocytes.
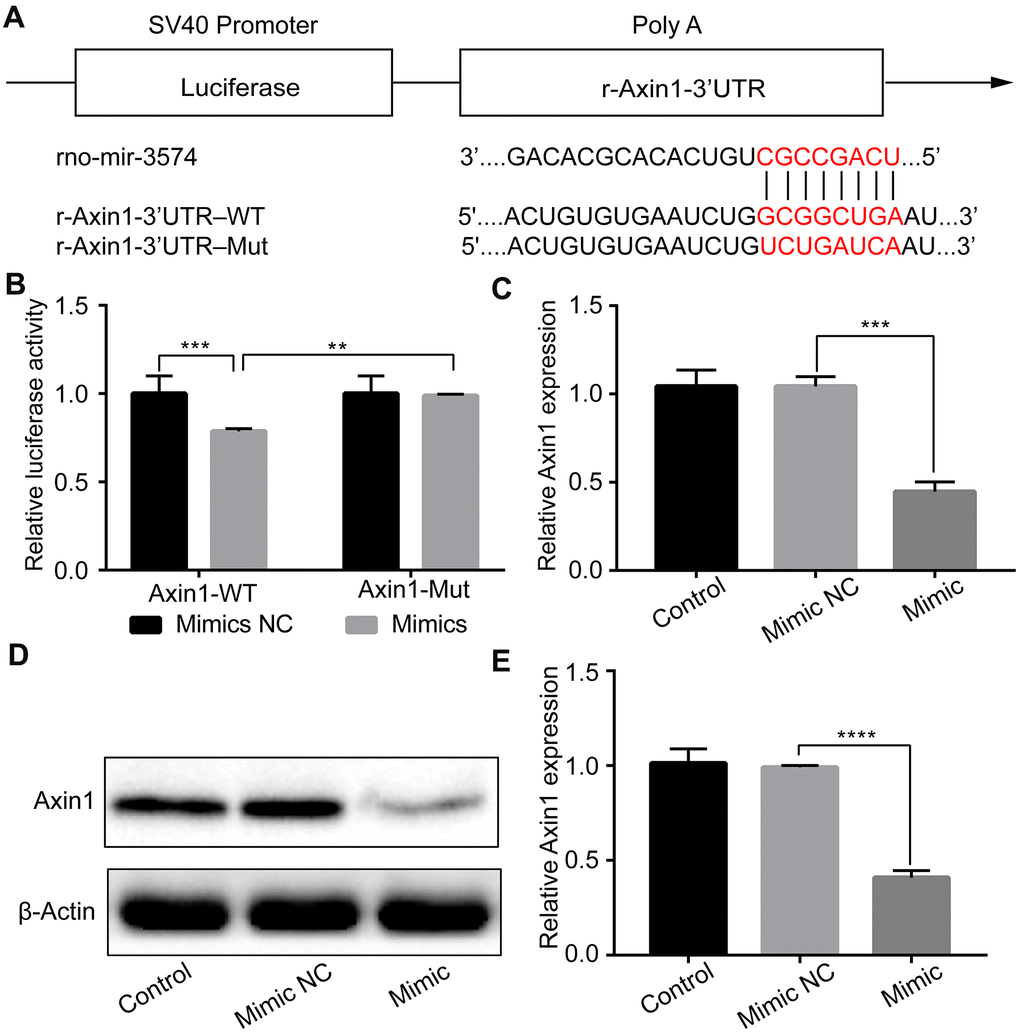
Figure 4. Axin1 is a target of miR-3574, and Axin1 could be inhibited by miR-3574 in H9c2 cells. (A) The potential binding site between miR-3574 and the 3’-UTR of Axin1. (B) Luciferase reporter assay was conducted by co-transfection of luciferase reporter containing 3’-UTR of Axin1 with miR-3574 mimic into 293T cells. (C–E) H9c2 cells were transfected with miR-3574 mimics or corresponding control. Expressions of Axin1 were examined by western blot analysis and RT-qPCR, respectively. NC: negative control. Data are represented as the mean ± SD from three independent experiments. * p < 0.05, ** p < 0.01.
Axin1 reverses the effects of miR-3574 overexpression on cell injury in IH-treated H9c2 cardiomyocytes
Finally, we performed rescue experiments to further explore whether Axin1 overexpression could rescue the anti-apoptosis function of miR-3574 in IH-induced cardiomyocytes. H9c2 cells was transfected with miR-3574 mimics together without or with pcDNA3.1- Axin1 under IH condition. The results showed that the reduced level of Axin1 caused by miR-3574 overexpression could be partly rescued by co-transfection with pcDNA3.1-Axin1 (P<0.05) (Figure 5B). At the same time, functional experiments further demonstrated that the reintroduction of Axin1 could partially reverse the effects of miR-3574 on the cell viability (P<0.05) (Figure 5A) and apoptosis (P<0.05) (Figure 5B) in the IH-treated H9c2 cardiomyocytes. Taken together, our findings indicated that Axin1 acts as a downstream target of miR-3574 in the determination of cell injury in IH-treated H9c2 cardiomyocytes.
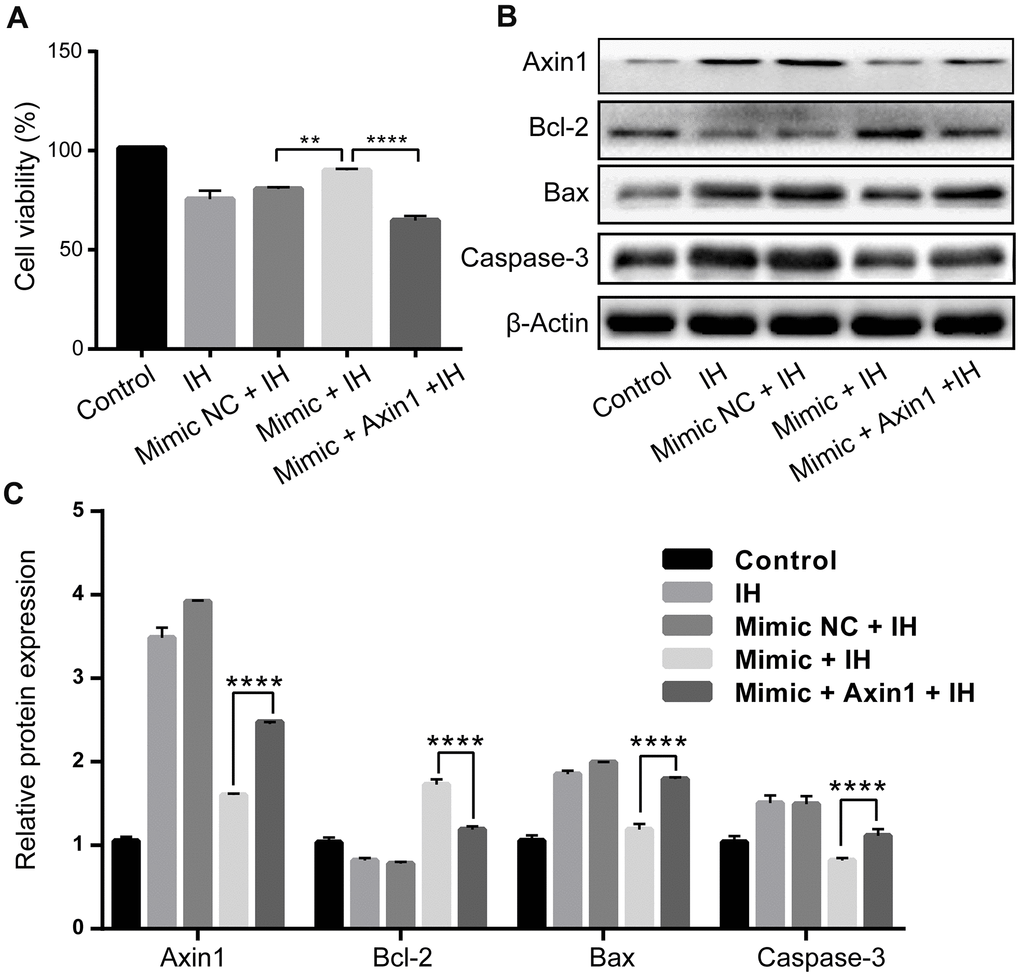
Figure 5. Upregulation of miR-3574 inhibits IH-induced H9c2 cardiomyocyte injury via Axin1. miR-3574 mimic, pcDNA3.1-Axin1 plasmid, and scrambled control were transfected into H9c2 cells. Cells without transfection were served as control. (A) Cell viability. (B, C) Western blot assays of Caspase-3, Bax, Bcl-2, and Axin1 protein. NC: negative control. IH: intermittent hypoxia. Data are represented as the mean ± SD from three independent experiments. * p < 0.05, ** p < 0.01.
Discussion
In the present study, we found that IH treatment induced cell injury and inhibited the expression of miR-3574 in the H9c2 cardiomyocytes. miR-3574 overexpression reversed the effects of IH treatment on the cell viability and apoptosis in H9c2 cells. Using bioinformatics prediction, Axin1 was explored to be one of the potential targets of miR-3574. Next, the Luciferase reporter assay confirmed that miR-3574 suppressed the expression of Axin1 by binding with the 3’UTR of Axin1. Furthermore, it was found that miR-3574 further suppressed the mRNA and protein levels of Axin1 in the H9c2 cardiomyocytes. More importantly, enforced expression of Axin1 could partly reverse the effects of miR-3574 overexpression on cell damage in the IH treated H9c2 cardiomyocytes. Our work first revealed that miR-3574 is a critical regulator in the myocardial IH injury, which may provide a potential therapeutic target in OSA-related heart disease.
There is increased recognition that OSA, characterized by intermittent hypoxia (IH), is deemed as a risk factor for the development of many cardiovascular diseases such as hypertension [4], myocardial ischemia [6], heart failure [7], atherosclerosis [17], and coronary artery disease [18]. The IH leads to a number of potential adverse consequences, including activation of inflammation [19], insulin resistance [20], oxidative stress [21], sympathetic activation [22], and endothelial dysfunction [23]. Thus, it contributes to OSA-associated cardiovascular pathologies. For example, previous studies showed that IH could activate the factors of NF-kappaB-dependent inflammation resulting in OSA- related cardiovascular dysfunction [24]. Meanwhile, it is reported that IH-induced oxidative stress appears to involve in the hypersensitivity to myocardial infarction [25]. Additionally, our previous study observed that IH also could cause endothelial dysfunction [26], which represents one of the initial steps of atherosclerosis development. In our study, we demonstrated that IH treatment markedly decreased cell viability and altered the expression levels of apoptosis-associated proteins in H9c2 cardiac myocytes. In the recent years, how to relieve IH-related cardiomyocytes injury has attracted increasing attention.
Accumulative data indicates that a number of miRNAs have taken part in the pathogenesis of cardiovascular diseases [27–29]. Moreover, increasing evidence confirmed that a large number of miRNAs could inhibit or promote cardiomyocytes apoptosis via mediating the downstream pathways and targets. For instance, Yang et al demonstrated that miR-320 participate in the cardioprotective effect of insulin against H9c2 cardiomyocytes apoptosis by targeting survivin [30]. At the same time, Pan et al found that miR-133b-5p contributes to HPC-induced cardioprotection by the inhibition of caspase-3 and caspase-8 apoptotic signaling [31]. In 2020, Zhang and coworkers found that miR-885 could attenuate hypoxia/reoxygenation-induced cell apoptosis via modulation of AKT/mTOR signaling in human cardiomyocytes [32]. As yet, very little is known about the function of miRNAs in OSA-related cardiovascular diseases. Our study first revealed that overexpression of miR-3574 could lead to a striking increase in cardiomyocyte viability and a remarkable decrease in H9c2 cell apoptosis following exposure to IH. That is to say, miR-3574 overexpression inhibits cardiomyocytes apoptosis under IH condition. Together, all these findings suggest that miR-3574 may have a pivotal role in the progression of IH cardiomyopathy. However, the underlying mechanisms remain unknown.
To further determine the underlying mechanism of miR-3574 in H9c2 cell viability and apoptosis under IH condition, we further carried out dual-luciferase reporter assay and bioinformatic analysis. We surprisingly found that the 3’-UTR of Axin1 contained the potential conserved binding sites of miR-3574. Axin1, a cytoplasmic protein, is an important regulator of the Wnt signaling pathway via inhibition of β-catenin [33, 34]. It was reported that Axin1 is a key mediator of cell apoptosis in human melanoma cells by inhibiting the BRAFV600E [35]. Thus, we hypothesized that Axin1involved in the process of miR-3574 regulating IH-induced H9c2 cell apoptosis. Indeed, we next demonstrated that miR-3574 overexpression was related to the inhibition of luciferase activity. In addition, we also observed that miR-3574 overexpression significantly reduced the mRNA and protein levels of Axin1. At last, rescue experiments further validated that re-introduction of Axin1 in IH-treatment of H9c2 cardiomyocytes transfected with miR-3574 mimics could significantly reversed the function of miR-3574 mimics. All of above results confirmed that Axin1 was a direct target of miR-3574, which could mediate the roles of miR-3574 in H9c2 cells viability and apoptosis under IH condition.
In our study, we first showed the negative correlation of miR-3574-Axin1 pair in vitro, and revealed that miR-3574 could alleviate IH-induced cardiomyocyte injury via inhibition of Axin1 in vitro. This result highlights the important role of miRNA regulation in OSA-related cardiovascular diseases. However, the study also harbors some limitations. Firstly, the morphological change of apoptosis was not investigated, such as the use of immunofluorescence and TUNEL assay. Secondly, this study is mainly focused their attention on only in vitro cell-based experiments. Thus, animal experiments on miR-3574 knock in/out should be conducted in future research. Fourthly, more markers indicating cardiomyocyte injury should be provided such as LDH, cTN, cleaved caspase-3, and so on. Finally, though the sequence in miR-3574 and Axin1 3’-UTR was highly conserved among multiple species, the function of miR-3574 and its clinical applicability are still needed to be further explored.
Conclusions
In conclusion, we first confirmed that miR-3574 overexpression inhibited IH-induced apoptosis in cardiomyocytes by targeting Axin1. Our findings identify that miR-3574 exerts a prominent cardioprotective effect on IH-induced cardiomyocyte injury, which seems to provide a novel target for treating OSA-related heart diseases.
Materials and Methods
Cell culture
H9c2 cardiomyocytes were obtained from Chinese Academy of Sciences Shanghai Cell Bank. Cells were routinely cultured in DMEM (HyClone) containing 10% fetal bovine serum (Gibco) in a 5% CO2 humidified incubator at 37° C.
IH treatment
When H9c2 cardiomyocytes reached 70-80% confluence, IH stimulation was carried out according to the method as we previously described [36]. Briefly, cells were first maintained under hypoxia for 35 min, which was induced by flushing a mixed air of 5% CO2, 1% O2 and 94% N2. Subsequently, cells were then cultured under normoxia condition for 25 min (5% CO2, 21% O2 balanced with N2). Repeated IH treatment was conducted for 6 cycles.
RNA isolation and qRT-PCR
Total RNA was extracted from H9c2 cardiomyocytes by use of the Trizol Reagent (Invitrogen, USA). The quality was determined by gel electrophoresis and NanoDrop 2000 (Thermo Fisher Scientific, USA). According to the manufacturers’ protocols, the reverse transcription of miRNA and mRNA from the total RNA was employed by using Mir-X™ miRNA First Strand Synthesis Kit (Clontech, USA) and PrimeScript™ RT Reagent Kit (Takara, China), respectively. qRT-PCR reactions were prepared by using SYBR Green PCR Master Mix (Thermo Fisher Scientific) and performed in ABI 7500 real-time RT-PCR system. β-actin and U6 were used as internal controls. All primers used in this study were listed in Table 1. Relative expression levels of miRNA and mRNA was calculated using the 2−∆∆Ct method. All experiments were conducted in triplicate.
Table 1. Primers used for RT-qPCR.
| ID | Sequence (5′-3′) |
| miR-3574 | Sense: CATCAGCCGCTGTCACACG |
| Antisense: CCAGTGCGTGTCGTGGAGT | |
| miR-3574 mimic | Sense: UCAGCCGCUGUCACACGCACAG |
| Antisense: CUGUGCGUGUGACAGCGGCUGA | |
| miR-3574 mimic NC | Sense: UUGUACUACACAAAAGUACUG |
| Antisense: GUACUUUUGUGUAGUACAAUU | |
| miR-3574 inhibitor | CUGUGCGUGUGACAGCGGCUGA |
| miR-3574 inhibitor NC | CAG UAC UUU UGU GUA GUA CAA |
| Axin1 | Sense: GAAGACGGCGATCCATCG |
| Antisense: GGATGCTCTCAGGGTTCT | |
| U6 | Sense: CTCGCTTCGGCAGCACA |
| Antisense: AACGCTTCACGAATTTGCGT | |
| β-Actin | Sense: CGAGTACAACCTTCTTGCAGC |
| Antisense: ACCCATACCCACCATCACAC |
Cell counting kit-8 (CCK-8) assay
CCK-8 assay was performed to evaluate cell viability. H9c2 cells were maintained in 96-well plates and gained the relevant treatments, including IH, transfection. For CCK8 detection, 10 μl/well of CCK-8 solution was added into each well. The absorbance was determined with the use of a microplate reader (Bio-Rad) at 450 nm.
Western blot analysis
Total proteins were first extracted from H9c2 cardiomyocytes using RIPA buffer (Beyotime, China). We used the BCA Protein Assay Kit (Thermo Scientific, USA) to detect the protein concentration. Subsequently, we separated equal amounts of protein on 10% SDS-PAGE, and then transferred the proteins to a PVDF membrane. After blocking with 5% nonfat milk, we then incubated the membrane with appropriate primary antibodies at 4° C overnight. Next, we employed TBST to wash the membrane three times. Then, it was incubated with secondary antibodies conjugated with horseradish peroxidase (Santa Cruz Biotechnology) for 1 h at room temperature. At last, the signals were visualized on X-ray films by using enhanced chemiluminescence (ECL). The bands were analyzed with the use of ImageJ software. The expression level of relative proteins was normalized to β-Actin.
Cell transfection
H9c2 cardiomyocytes were detached with trypsin (Gibco) and counted the day before transfection. H9c2 cardiomyocytes were cultured to reach 90% confluence on the day of transfection. The miR-3574 inhibitor and mimic, and scrambled control were obtained from Sangon Biotech Co. (Shanghai, China), which were listed in Table 1. At the same time, the pcDNA3.1-Axin1 plasmid and vector were also obtained from Sangon Biotech Co. Then, we transfected miRNAs and pcDNA3.1 plasmids into H9c2 cells with Lipofectamine 3000 (Invitrogen, USA). All transfections were carried out before the IH treatment, and then cultured for 48 h for subsequent experimentation.
Dual-luciferase reporter assay
We generated the mutant 3’-UTR of Axin1 by use of the Quick-Change Site-Directed Mutagenesis kit (Stratagene, USA). The 3’-UTR of Axin1 contained either the miR-3574 binding site or the mutant 3’-UTR of Axin1, which was cloned into the psiCHECK-2 vectors (Promega, USA) to synthesize the psiCHECK-Axin1 3’-UTR Luciferase reporter plasmid. Following the manufacturer’s instruction, luciferase reporter plasmids and miR-3574 mimic or miR-NC were then co-transfected into H9c2 with the help of Lipofectamine 3000 reagent (Invitrogen, USA). After 48 h, we applied a Dual-Luciferase reporter gene assay kit (Promega, USA) to detect the Luciferase activities. Renilla luciferase was served as normalisation. The experiment was repeated three times.
Statistical analysis
All data were displayed as the mean ± SD from at least three independent experiments. GraphPad Prism 7 and SPSS 22.0 were used to analyze the data. We analyzed the differences between two independent groups by using Student’s t tests. One-way ANOVA were used for the comparison of differences among multiple groups. P < 0.05 was deemed statistically significant.
Author Contributions
Qingshi Chen and Guofu Lin: carried out the qRT-PCR, dual-luciferase reporter assay and western blot analysis, conceived of the study, and wrote this manuscript. Yongfa Chen, Chaowei Li and Lizhen Wu: performed the statistical analysis and participated in the design of the study. Xin Hu: carried out the CCK-8 assay, cell transfection, cell culture, and IH treatment. Qichang Lin: conceived of the study.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Funding
This study was funded by the National Natural Science Foundation of China [grant number: 82000094, 81370182 and 81870074]; Science and Technology Projects of Quanzhou [grant numbers: 2018N007S]; Science and Technology Project of Fujian Education Department [grant number: JT180199] and Natural Science Foundation of Fujian Province [grant number: 2020J01203].
References
- 1. Franczak A, Bil-Lula I, Sawicki G, Fenton M, Ayas N, Skomro R. Matrix metalloproteinases as possible biomarkers of obstructive sleep apnea severity - a systematic review. Sleep Med Rev. 2019; 46:9–16. https://doi.org/10.1016/j.smrv.2019.03.010 [PubMed]
- 2. Boulet L, Flore P, Le Gouellec A, Toussaint B, Pépin JL, Faure P. Is tryptophan metabolism involved in sleep apnea-related cardiovascular co-morbidities and cancer progression? Med Hypotheses. 2015; 85:415–23. https://doi.org/10.1016/j.mehy.2015.06.022 [PubMed]
- 3. Ozeke O, Ozer C, Gungor M, Celenk MK, Dincer H, Ilicin G. Chronic intermittent hypoxia caused by obstructive sleep apnea may play an important role in explaining the morbidity-mortality paradox of obesity. Med Hypotheses. 2011; 76:61–63. https://doi.org/10.1016/j.mehy.2010.08.030 [PubMed]
- 4. André S, Andreozzi F, Van Overstraeten C, Ben Youssef S, Bold I, Carlier S, Gruwez A, Bruyneel AV, Bruyneel M. Cardiometabolic comorbidities in obstructive sleep apnea patients are related to disease severity, nocturnal hypoxemia, and decreased sleep quality. Respir Res. 2020; 21:35. https://doi.org/10.1186/s12931-020-1284-7 [PubMed]
- 5. Huang B, Liu H, Scherlag BJ, Sun L, Xing S, Xu J, Luo M, Guo Y, Cao G, Jiang H. Atrial fibrillation in obstructive sleep apnea: neural mechanisms and emerging therapies. Trends Cardiovasc Med. 2020. [Epub ahead of print]. https://doi.org/10.1016/j.tcm.2020.01.006 [PubMed]
- 6. Konishi T, Kashiwagi Y, Funayama N, Yamamoto T, Murakami H, Hotta D, Tanaka S. Obstructive sleep apnea is associated with increased coronary plaque instability: an optical frequency domain imaging study. Heart Vessels. 2019; 34:1266–79. https://doi.org/10.1007/s00380-019-01363-8 [PubMed]
- 7. Zhan X, Li L, Wu C, Chitguppi C, Huntley C, Fang F, Wei Y. Effect of uvulopalatopharyngoplasty (UPPP) on atherosclerosis and cardiac functioning in obstructive sleep apnea patients. Acta Otolaryngol. 2019; 139:793–97. https://doi.org/10.1080/00016489.2019.1633475 [PubMed]
- 8. Liu Q, Wu DH, Han L, Deng JW, Zhou L, He R, Lu CJ, Mi QS. Roles of microRNAs in psoriasis: immunological functions and potential biomarkers. Exp Dermatol. 2017; 26:359–67. https://doi.org/10.1111/exd.13249 [PubMed]
- 9. Hashimoto-Gotoh A, Kitao K, Miyazawa T. Persistent infection of simian foamy virus derived from the Japanese macaque leads to the high-level expression of microRNA that resembles the miR-1 microRNA precursor family. Microbes Environ. 2020; 35:ME19130. https://doi.org/10.1264/jsme2.ME19130 [PubMed]
- 10. Chandan K, Gupta M, Sarwat M. Role of host and pathogen-derived MicroRNAs in immune regulation during infectious and inflammatory diseases. Front Immunol. 2020; 10:3081. https://doi.org/10.3389/fimmu.2019.03081 [PubMed]
- 11. Li Y, Liu X, Yin Z, You Y, Zou Y, Liu M, He Y, Zhang H, Zheng X, Zhang Z, Wang P. MicroRNA-like milR236, regulated by transcription factor MoMsn2, targets histone acetyltransferase MoHat1 to play a role in appressorium formation and virulence of the rice blast fungus magnaporthe oryzae. Fungal Genet Biol. 2020; 137:103349. https://doi.org/10.1016/j.fgb.2020.103349 [PubMed]
- 12. Zhang J, Wei X, Zhang W, Wang F, Li Q. MiR-326 targets MDK to regulate the progression of cardiac hypertrophy through blocking JAK/STAT and MAPK signaling pathways. Eur J Pharmacol. 2020; 872:172941. https://doi.org/10.1016/j.ejphar.2020.172941 [PubMed]
- 13. Foinquinos A, Batkai S, Genschel C, Viereck J, Rump S, Gyöngyösi M, Traxler D, Riesenhuber M, Spannbauer A, Lukovic D, Weber N, Zlabinger K, Hašimbegović E, et al. Preclinical development of a miR-132 inhibitor for heart failure treatment. Nat Commun. 2020; 11:633. https://doi.org/10.1038/s41467-020-14349-2 [PubMed]
- 14. Gao F, Kataoka M, Liu N, Liang T, Huang ZP, Gu F, Ding J, Liu J, Zhang F, Ma Q, Wang Y, Zhang M, Hu X, et al. Therapeutic role of miR-19a/19b in cardiac regeneration and protection from myocardial infarction. Nat Commun. 2019; 10:1802. https://doi.org/10.1038/s41467-019-09530-1 [PubMed]
- 15. Yang X, Niu X, Xiao Y, Lin K, Chen X. MiRNA expression profiles in healthy OSAHS and OSAHS with arterial hypertension: potential diagnostic and early warning markers. Respir Res. 2018; 19:194. https://doi.org/10.1186/s12931-018-0894-9 [PubMed]
- 16. Khalyfa A, Zhang C, Khalyfa AA, Foster GE, Beaudin AE, Andrade J, Hanly PJ, Poulin MJ, Gozal D. Effect on intermittent hypoxia on plasma exosomal micro RNA signature and endothelial function in healthy adults. Sleep. 2016; 39:2077–90. https://doi.org/10.5665/sleep.6302 [PubMed]
- 17. Kim S, Lee KY, Kim NH, Abbott RD, Kim C, Lee SK, Kim SH, Shin C. Relationship of obstructive sleep apnoea severity and subclinical systemic atherosclerosis. Eur Respir J. 2020; 55:1900959. https://doi.org/10.1183/13993003.00959-2019 [PubMed]
- 18. Glantz H, Thunström E, Johansson MC, Wallentin Guron C, Uzel H, Ejdebäck J, Nasic S, Peker Y. Obstructive sleep apnea is independently associated with worse diastolic function in coronary artery disease. Sleep Med. 2015; 16:160–67. https://doi.org/10.1016/j.sleep.2014.08.018 [PubMed]
- 19. Chen JK, Guo MK, Bai XH, Chen LQ, Su SM, Li L, Li JQ. Astragaloside IV ameliorates intermittent hypoxia-induced inflammatory dysfunction by suppressing MAPK/NF-κB signalling pathways in Beas-2B cells. Sleep Breath. 2020; 24:1237–45. https://doi.org/10.1007/s11325-019-01947-8 [PubMed]
- 20. Murphy AM, Thomas A, Crinion SJ, Kent BD, Tambuwala MM, Fabre A, Pepin JL, Roche HM, Arnaud C, Ryan S. Intermittent hypoxia in obstructive sleep apnoea mediates insulin resistance through adipose tissue inflammation. Eur Respir J. 2017; 49:1601731. https://doi.org/10.1183/13993003.01731-2016 [PubMed]
- 21. Badran M, Yassin BA, Lin DT, Kobor MS, Ayas N, Laher I. Gestational intermittent hypoxia induces endothelial dysfunction, reduces perivascular adiponectin and causes epigenetic changes in adult male offspring. J Physiol. 2019; 597:5349–64. https://doi.org/10.1113/JP277936 [PubMed]
- 22. Kimura H, Ota H, Kimura Y, Takasawa S. Effects of intermittent hypoxia on pulmonary vascular and systemic diseases. Int J Environ Res Public Health. 2019; 16:3101. https://doi.org/10.3390/ijerph16173101 [PubMed]
- 23. Badran M, Abuyassin B, Ayas N, Laher I. Intermittent hypoxia impairs uterine artery function in pregnant mice. J Physiol. 2019; 597:2639–50. https://doi.org/10.1113/JP277775 [PubMed]
- 24. Xu XM, Yao D, Cai XD, Ding C, Lin QD, Wang LX, Huang XY. Effect of chronic continual- and intermittent hypoxia-induced systemic inflammation on the cardiovascular system in rats. Sleep Breath. 2015; 19:677–84. https://doi.org/10.1007/s11325-014-1075-9 [PubMed]
- 25. Totoson P, Fhayli W, Faury G, Korichneva I, Cachot S, Baldazza M, Ribuot C, Pépin JL, Lévy P, Joyeux-Faure M. Atorvastatin protects against deleterious cardiovascular consequences induced by chronic intermittent hypoxia. Exp Biol Med (Maywood). 2013; 238:223–32. https://doi.org/10.1177/1535370212473696 [PubMed]
- 26. Chen Q, Lin G, Huang J, Chen L, Liu Y, Huang J, Zhang S, Lin Q. Inhibition of miR-193a-3p protects human umbilical vein endothelial cells against intermittent hypoxia-induced endothelial injury by targeting FAIM2. Aging (Albany NY). 2020; 12:1899–909. https://doi.org/10.18632/aging.102729 [PubMed]
- 27. Kir D, Schnettler E, Modi S, Ramakrishnan S. Regulation of angiogenesis by microRNAs in cardiovascular diseases. Angiogenesis. 2018; 21:699–710. https://doi.org/10.1007/s10456-018-9632-7 [PubMed]
- 28. Zhou SS, Jin JP, Wang JQ, Zhang ZG, Freedman JH, Zheng Y, Cai L. miRNAS in cardiovascular diseases: potential biomarkers, therapeutic targets and challenges. Acta Pharmacol Sin. 2018; 39:1073–84. https://doi.org/10.1038/aps.2018.30 [PubMed]
- 29. Gong YY, Luo JY, Wang L, Huang Y. MicroRNAs regulating reactive oxygen species in cardiovascular diseases. Antioxid Redox Signal. 2018; 29:1092–107. https://doi.org/10.1089/ars.2017.7328 [PubMed]
- 30. Yang N, Wu L, Zhao Y, Zou N, Liu C. MicroRNA-320 involves in the cardioprotective effect of insulin against myocardial ischemia by targeting survivin. Cell Biochem Funct. 2018; 36:166–71. https://doi.org/10.1002/cbf.3328 [PubMed]
- 31. Pan YL, Han ZY, He SF, Yang W, Cheng J, Zhang Y, Chen ZW. miR-133b-5p contributes to hypoxic preconditioning-mediated cardioprotection by inhibiting the activation of caspase-8 and caspase-3 in cardiomyocytes. Mol Med Rep. 2018; 17:7097–104. https://doi.org/10.3892/mmr.2018.8784 [PubMed]
- 32. Meng X, Mei L, Zhao C, Chen W, Zhang N. miR-885 mediated cardioprotection against hypoxia/reoxygenation-induced apoptosis in human cardiomyocytes via inhibition of PTEN and BCL2L11 and modulation of AKT/mTOR signaling. J Cell Physiol. 2020; 235:8048–57. https://doi.org/10.1002/jcp.29460 [PubMed]
- 33. Sue Ng S, Mahmoudi T, Li VS, Hatzis P, Boersema PJ, Mohammed S, Heck AJ, Clevers H. MAP3K1 functionally interacts with Axin1 in the canonical Wnt signalling pathway. Biol Chem. 2010; 391:171–80. https://doi.org/10.1515/BC.2010.028 [PubMed]
- 34. Zhou Y, Shu B, Xie R, Huang J, Zheng L, Zhou X, Xiao G, Zhao L, Chen D. Deletion of Axin1 in condylar chondrocytes leads to osteoarthritis-like phenotype in temporomandibular joint via activation of β-catenin and FGF signaling. J Cell Physiol. 2019; 234:1720–29. https://doi.org/10.1002/jcp.27043 [PubMed]
- 35. Biechele TL, Kulikauskas RM, Toroni RA, Lucero OM, Swift RD, James RG, Robin NC, Dawson DW, Moon RT, Chien AJ. Wnt/β-catenin signaling and AXIN1 regulate apoptosis triggered by inhibition of the mutant kinase BRAFV600E in human melanoma. Sci Signal. 2012; 5:ra3. https://doi.org/10.1126/scisignal.2002274 [PubMed]
- 36. Lin G, Huang J, Chen Q, Chen L, Feng D, Zhang S, Huang X, Huang Y, Lin Q. miR-146a-5p mediates intermittent hypoxia-induced injury in H9c2 cells by targeting XIAP. Oxid Med Cell Longev. 2019; 2019:6581217. https://doi.org/10.1155/2019/6581217 [PubMed]