



Introduction
Bladder urothelial cancer (BUC) is one of the most common genitourinary carcinomas in China [1]. It is also a significant cause of mortality in Western countries [2, 3]. BUC is categorized as muscle-invasive bladder cancer (MIBC) and non–muscle-invasive bladder cancer (NMIBC). Despite advances in surgical techniques and the development of novel drugs, NMIBC (stage Ta-1) with high morbidity tends to recur, whereas MIBC (stage T2-4) tends to metastasize within 2 years [3, 4]. Thus, it is important to identify the molecular mechanisms underlying tumorigenesis and metastasis of BUC so that new therapeutic targets and modalities can be developed to prevent and treat BUC [5]. BUC is thought to arise as a result of an accumulation of genetic alterations. At present, most studies on the etiology of BUC have focused on genetic transformation. Many oncogenes, such as C-myc [6], Bcl-2 [7], FGFR3 [8], and c-erbB-2 [9], have been found to be associated with BUC prognosis and progression [10]. However, the development of BUC is a complicated pathological process that requires further study.
Protein arginine methyltransferase 5 (PRMT5) is the predominant type II methyltransferase and is involved in numerous cellular and biological processes, such as Golgi homeostasis, nucleic acid metabolism, transcriptional regulation, cell cycle regulation, and ribosome biosynthesis [11, 12]. PRMT5 is a multifunctional gene with important functions in cells. Recent reports indicate that PRMT5 is overexpressed in a variety of human malignant tumors, suggesting that PRMT5 overexpression is an important high-risk factor and determinant of tumor properties. PRMT5 has been found to be upregulated in malignant tumors, such as gastric, lung, and prostate cancers [13–16]. One study demonstrated that overexpression and increased intranuclear accumulation of PRMT5 through methylation of Zn-finger protein were associated with a higher risk of defects in alternative splicing, leading to immortalized breast epithelial cells in humans [17]. The PRMT5 gene is a key regulator of amplification, migration, and metastasis of malignant tumors and participates in regulation of the cell cycle and autophagy. However, evidence is lacking regarding the relationship between PRMT5 expression and BUC.
Few diagnostic and prognostic genetic markers have been verified in BUC. Based on previous studies, this study was designed to detect PRMT5 gene expression in BUC and to assess its clinical significance. In addition, we studied the effect of PRMT5 on the proliferation and aggression of BUC.
Results
PRMT5 is upregulated in BUC
We performed bioinformatics analysis in three BUC data sets from the Oncomine database and found that PRMT5 was upregulated in superficial or infiltrating BUC tissue compared with nontumor bladder tissues (Figure 1A–1D). On further bioinformatics analysis using The Cancer Genome Atlas (TCGA) database, we confirmed that PRMT5 expression was higher in BUC tissue than in matched nonneoplastic bladder tissue (Figure 1E). In addition, in the TCGA database, high PRMT5 expression was associated with poor overall and progression-free survival in BUC patients (Figure 1F, 1G). To verify the database results, we determined the expression of PRMT5 mRNA in a cohort of 132 pairs of BUC and adjacent nontumor bladder tissues using quantitative and real-time polymerase chain reaction (qRT-PCR). We found that PRMT5 levels were significantly higher in BUC samples than in normal control samples (Figure 2A). In addition, PRMT5 protein was found to be overexpressed in BUC samples on western blotting assay and immunohistochemistry (Figure 2B–2D). Thus, we next focused on exploring the potential role of PRMT5 in BUC.
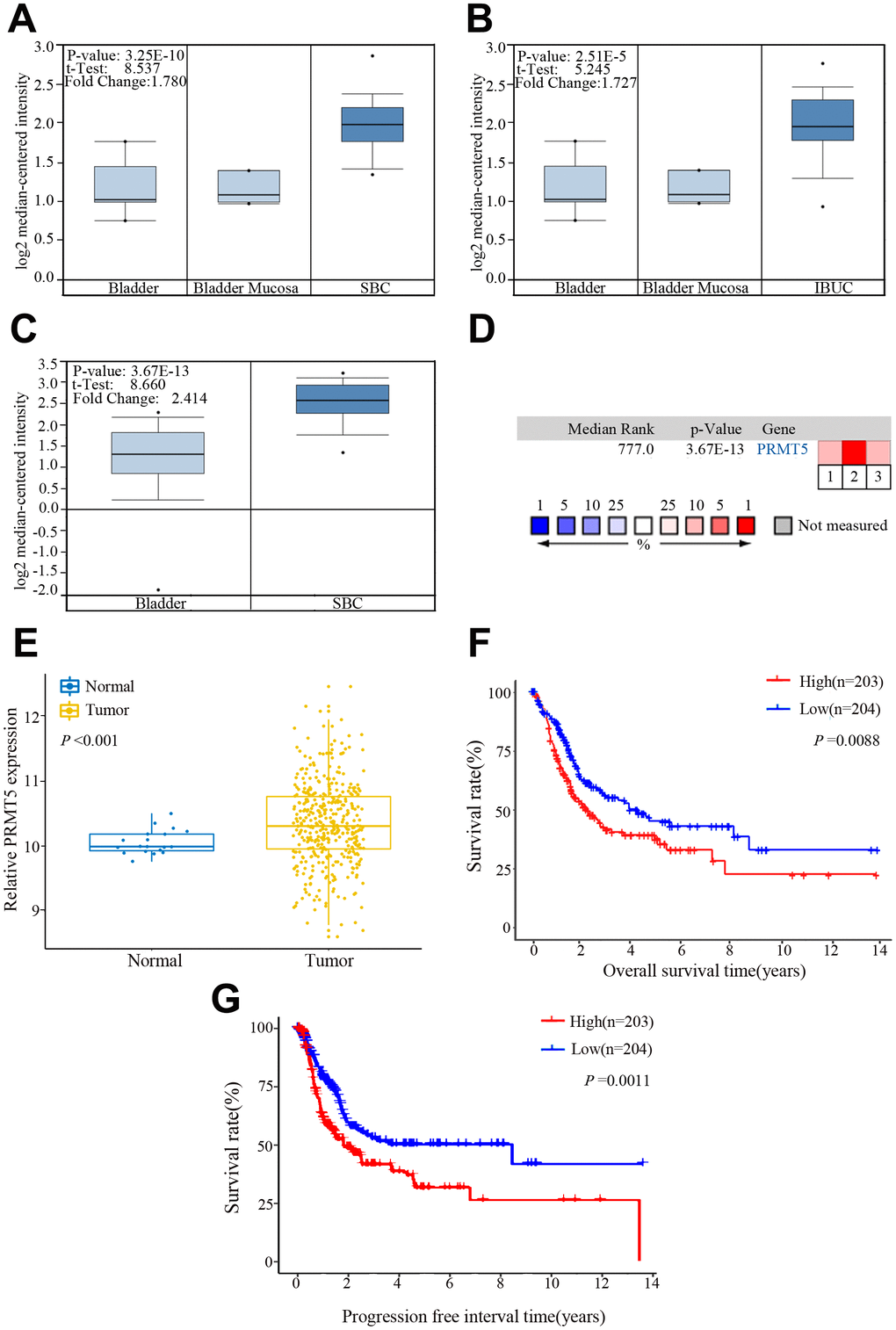
Figure 1. PRMT5 expression in bladder cancer in the Oncomine and The Cancer Genome Atlas (TCGA) databases. (A–C) Comparison of the PRMT5 expression levels in the Dyrskjot Bladder 3 and Sanchez-Carbayo Bladder 2 data sets of the Oncomine database. The threshold: P value: 1E-4, fold change: 1.5, gene rank: 10%. (D) A median-ranked analysis of the Dyrskjot Bladder 3 (1, 2) and Sanchez-Carbayo Bladder 2 (3) data sets from the Oncomine database. The colored squares revealed the median rank for PRMT5 across the three analyses (vs normal tissue). (E) Comparison of the PRMT5 expression level in bladder cancer and the normal tissue from the TCGA database. (F, G) Overall and progression-free survival times in bladder cancer patients with low versus high expression of PRMT5 assessed by Kaplan-Meier analysis from the TCGA cohorts. SBC: superficial bladder cancer, IBUC: infiltrating bladder urothelial carcinoma.
PRMT5 upregulation is correlated with poor prognosis in BUC patients
Figure 2E shows that patients with high PRMT5 expression had a worse prognosis compared with patients with low expression (5-year overall survival rates, 33.3% vs 58.2%, respectively; P = 0.0106). The Kaplan-Meier curves also demonstrate poorer overall survival of patients with high PRMT5 expression, compared with those with low expression, with MIBC (T2-4) (P = 0.0360), absence of lymph node metastasis (P = 0.0298), and high-grade tumors (P = 0.0426; Figure 2F–2H). However, there was no significant association between PRMT5 expression and clinicopathologic parameters in BUC patients (Table 1). In addition, multivariate Cox proportional hazards regression analysis demonstrated PRMT5 upregulation to be an independent prognostic risk factor for worse survival of BUC patients (P = 0.012, Table 2). Thus, PRMT5 upregulation is associated with poor prognosis in BUC.
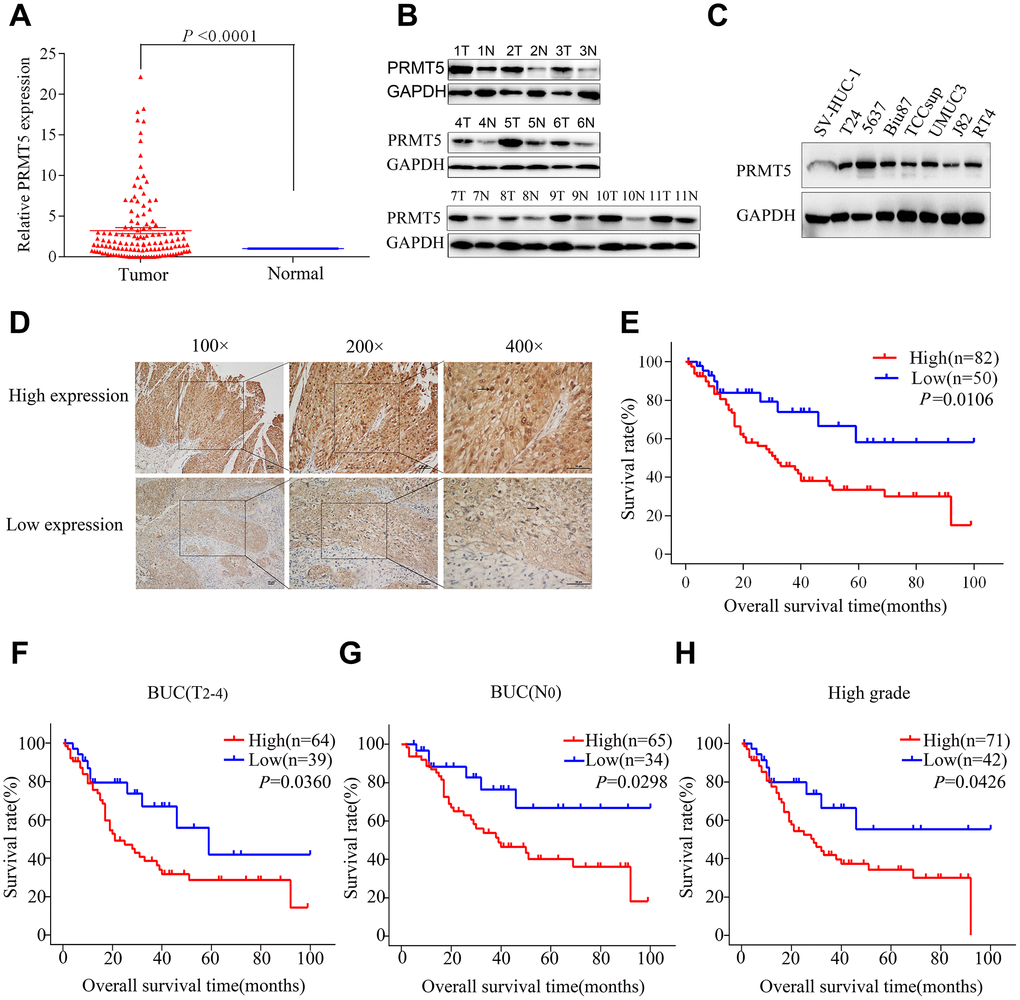
Figure 2. PRMT5 was upregulated and demonstrated prognostic significance in bladder cancer. (A) PRMT5 mRNA expression was significantly upregulated in bladder cancer tissue compared with that in adjacent normal tissues via qRT-PCR. (B) The PRMT5 protein level was upregulated in 11 pairs of bladder cancer tissues. (C) PRMT5 expression was upregulated in bladder cancer cell lines compared with immortalized human bladder epithelial SV-HUC-1 cells. (D) Representative images of immunohistochemistry of PRMT5 in bladder cancer tissues. (E) The Kaplan-Meier curve was applied to the survival analysis of bladder cancer patients with different PRMT5 expression levels from SYSUCC cohorts. (F–H) Positive correlation between overall survival and different PRMT5 expression levels from SYSUCC bladder cancer patients with muscle-invasive bladder cancer (F), absence of lymph node metastasis (G), and high-grade tumors (H). SYSUCC: Sun Yat-Sen University Cancer Center.
Table 1. The relationship between PRMT5 expression and clinicopathological characteristics in bladder cancer.
| Variables | No. | Expression of PRMT5 Level in BUC | χ2 | Pvalue | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Low | High | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Age (years) | 0.158 | 0.691 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| <65 | 71 | 28 | 43 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ≥65 | 61 | 22 | 39 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Gender | 0.789 | 0.374 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Male | 108 | 39 | 69 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Female | 24 | 11 | 13 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Tumor size (cm) | 0.002 | 0.961 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| <3 | 69 | 26 | 43 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ≥3 | 63 | 24 | 39 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| T classification | 2.580 | 0.630 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ta | 6 | 3 | 3 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| T1 | 22 | 7 | 15 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| T2 | 25 | 8 | 17 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| T3 | 44 | 20 | 24 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| T4 | 34 | 11 | 23 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| N classification | 0.005 | 0.946 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Negative | 84 | 32 | 52 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Positive | 48 | 18 | 30 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Grade | 0.168 | 0.681 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Low | 19 | 8 | 11 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| High/intermediate | 113 | 42 | 71 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Statistically significant (P < 0.05). BUC: bladder urothelial cancer, T and N classification: TNM stage. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Table 2. Univariate and multivariate analyses of clinicopathological characteristics for survival in patients with bladder cancer.
| Variables | Univariate analysis P value | Multivariate analysis | P value | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HR (95% CI) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Expression of PRMT5 | 0.014 | 2.434 (1.215-4.876) | 0.012 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Low | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| High | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Age | 0.081 | 1.542 (0.896-2.653) | 0.118 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| <65 years | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ≥65 years | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Gender | 0.130 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Male | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Female | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Tumor size | 0.169 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| <3 cm | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ≥3 cm | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| T classification | <0.001 | 1.576 (1.155-2.151) | 0.004 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ta | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| T1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| T2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| T3 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| T4 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| N classification | 0.001 | 1.482 (0.797-2.755) | 0.213 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Negative | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Positive | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Grade | 0.097 | 1.209 (0.536-2.727) | 0.674 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Low | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| High/intermediate | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HR: hazard ratio, CI: confidence interval. Bold values are statistically significant (P < 0.05). | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
PRMT5 promotes proliferation, migration, and invasion of BUC cells
We investigated the function of PRMT5 in BUC cells in vitro using western blotting and confirmed that the relative level of PRMT5 expression was downregulated in Biu87 and T24 cells by two specific siRNAs compared with that in the negative control group (Figure 3A). Cell proliferation was inhibited in cells with knockdown of PRMT5 as a result of siRNA. EdU assay was applied to explore the function of PRMT5 in promoting cell growth. There were significantly more EdU-positive T24 or Biu87 cells in the negative group than in the si-PRMT5 group after transfection of the indicated siRNA (Figure 3B). Next, the cell growth assay using cell counting kit-8 revealed that PRMT5 knockdown significantly decreased the number of the two indicated BUC cell lines (P < 0.05, Figure 3C). In the colony formation assay, both T24-siRNA and Biu87-siRNA cells formed fewer and smaller colonies than the negative control cells (P < 0.05, Figure 3D). Similarly, gene silencing of PRMT5 also significantly reduced BUC cell invasion and migration abilities (P < 0.05, Figure 3E, 3F). When we upregulated PRMT5 in the TCCsup cells (Figure 4A), the transfected TCCsup-PRMT5 cells demonstrated a significantly higher proliferative capacity compared with the respective control cells (P < 0.05, Figure 4B, 4C). In addition, the transwell migration assay and the cell scratch assay, which were performed to evaluate the invasion and migration ability of the transfected TCCsup-PRMT5 cells, indicated that PRMT5 significantly enhanced the migration capacity of BUC cells in vitro (P < 0.05, Figure 4D, 4E). These results suggest that PRMT5 promotes cell proliferation, invasion, and migration in BUC in vitro.
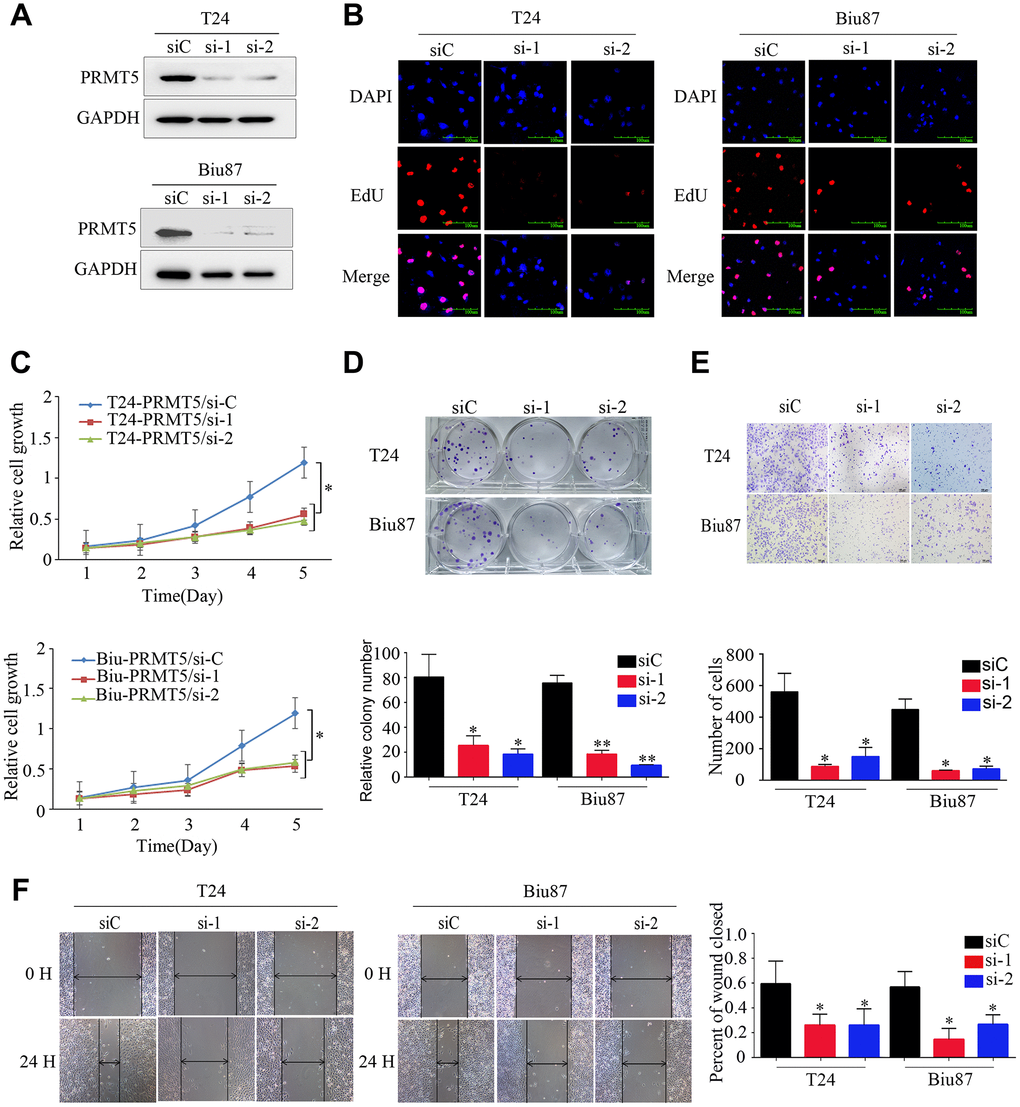
Figure 3. Downregulated expression of PRMT5 inhibited the proliferation and aggressiveness of bladder cancer cells. (A) PRMT5 was efficiently knocked down in T24-siRNA and Biu87-siRNA cells. (B–D) Cell proliferation was determined by EdU assay, CCK-8 assay, and colony formation assay of bladder cancer cells. (E, F) Cell invasion and migration capacities were determined using the transwell invasion assay and wound-healing assay for the indicated bladder cancer cells. Error bars show the standard error of the mean. siC: PRMT5-siRNA/negative control; si-1: PRMT5-siRNA/#1; si-2: PRMT5-siRNA/#2. *P < 0.05, **P < 0.01.
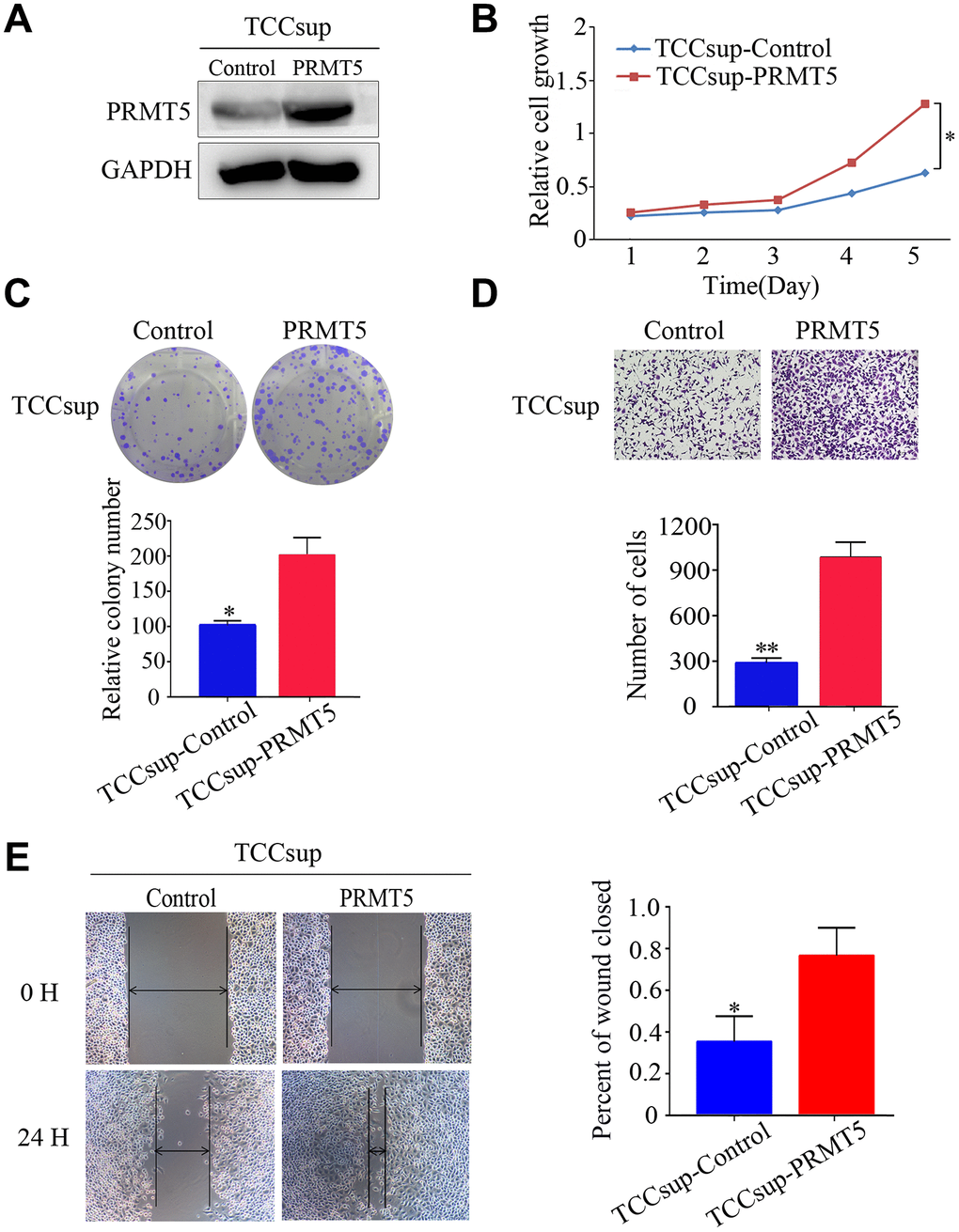
Figure 4. Upregulated expression of PRMT5 promoted the proliferation and aggressiveness of bladder cancer cells. (A) Western blotting showed successful overexpression of PRMT5 in TCCsup cells. (B, C) Cell proliferation was promoted by the CCK-8 assay and the colony formation assay. (D, E) Cell invasion and migration of bladder cancer cells were detected using the transwell assay and the wound-healing assay. Error bars show the standard error of the mean. *P < 0.05, **P < 0.01.
To confirm the effect of PRMT5 on tumor growth in vivo, a nude mouse subcutaneous xenograft tumor model was established. Western blotting identified the shRNA efficiency of PRMT5 downregulation at the protein level (Figure 5A). As shown in Figure 5B, the subcutaneous xenografts in mice inoculated with T24-sh1 cells were smaller than those in the control group. This result was confirmed using hematoxylin and eosin staining. The weight and size of the tumors were significantly reduced in the T24-shRNA group compared with the negative control group (P < 0.05, Figure 5C–5E).
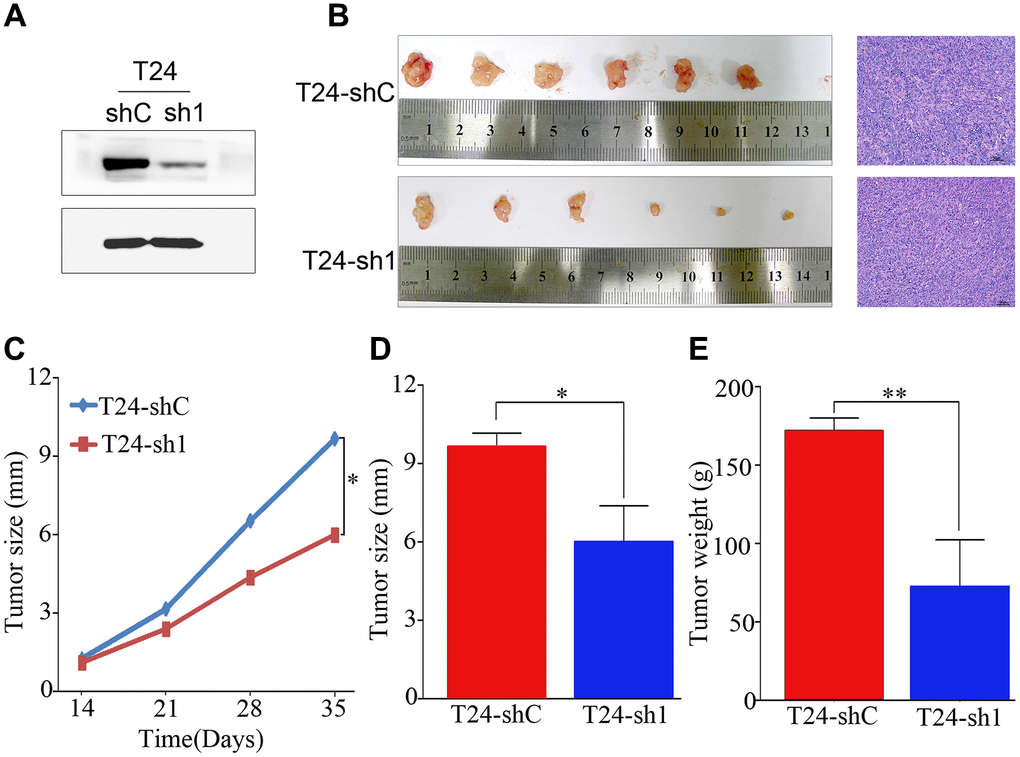
Figure 5. PRMT5 had a strong oncogenic function in bladder cancer. (A) Western blotting showed that PRMT5 was efficiently knocked down by PRMT5-shRNA. (B) Representative images of the tumor xenografts inoculated with T24-shC and T24-sh1 cells and hematoxylin and eosin staining of tumor xenografts are shown. (C–E) Growth curve and histogram analysis of the size and weight of xenograft tumors. Error bars show the standard error of the mean. sh, short hairpin RNA; T24-shC: T24-shRNA/negative control; T24-sh1: T24-shRNA. *P < 0.05, **P < 0.01.
PRMT5 affects cell cycle G1/S arrest and mTOR signaling pathway activity
To investigate the biological pathway of PRMT5 in relation to BUC pathogenesis, gene set enrichment analysis (GSEA) was conducted using GSE133624. GSEA enrichment plots show a significant positive correlation between cell cycle and mTOR signaling pathway and PRMT5 in BUC (Figure 6A, 6B). To confirm that PRMT5 promotes cell proliferation by regulating the cell cycle, flow cytometry was performed to determine the effect of PRMT5 on cell cycle G1/S arrest. The proportion of cells in the G0/G1 phase significantly decreased after treatment with PRMT5 knockdown. This arrest corresponded with the observed result of proliferation and self-renewal, as shown in Figure 3. Furthermore, the G0/G1 phase increased in response to PRMT5 overexpression in BUC cells (P < 0.01, Figure 6C–6F).
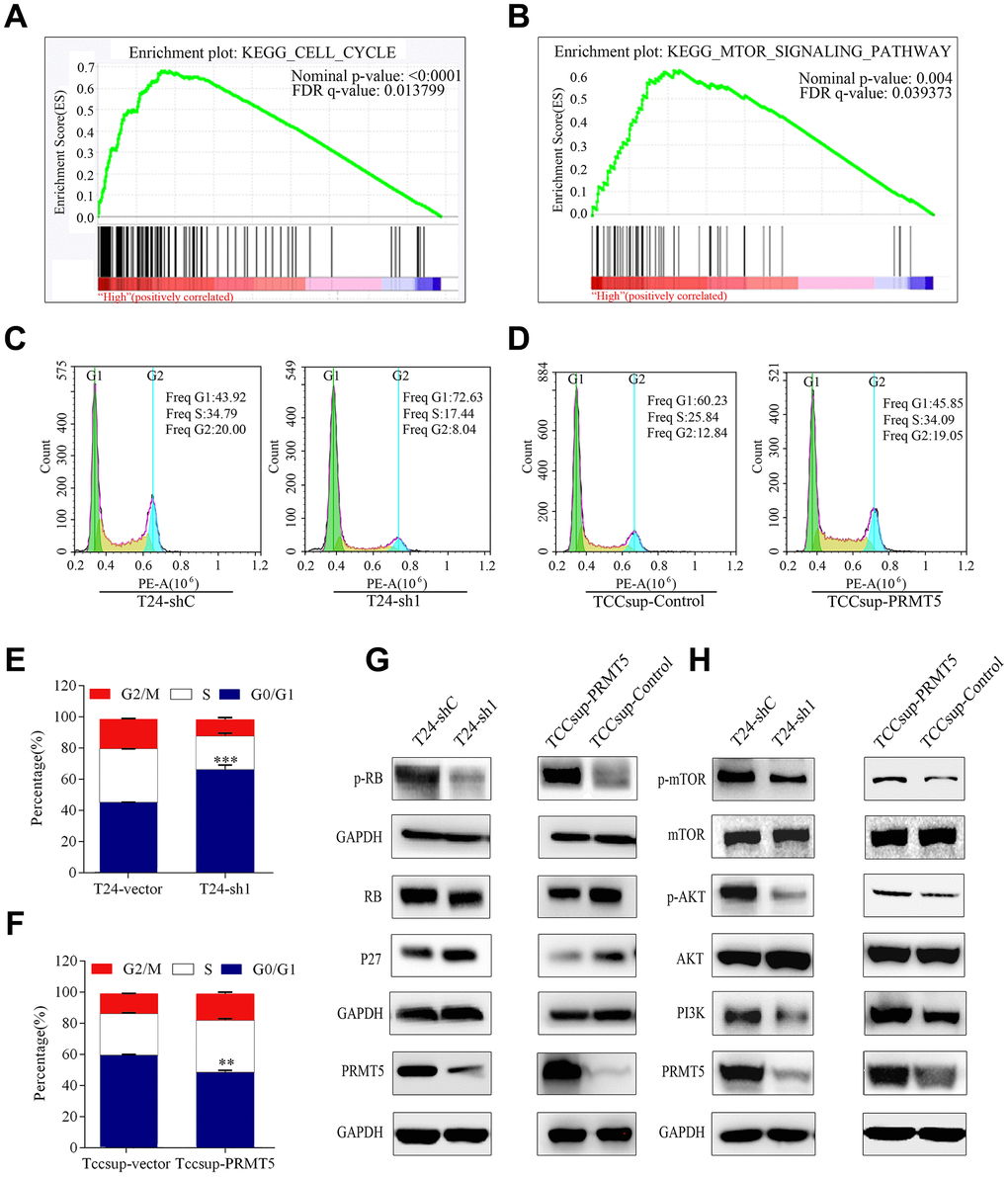
Figure 6. High PRMT5 expression in bladder cancer promoted tumor progression through the cell cycle and the mTOR signaling pathway. (A, B) GSEA indicated that high or low PRMT5 expression in bladder cancer is correlated with the cell cycle and the mTOR signaling pathway. Values were row-scaled to show the relative expression. (C) Flow cytometric analysis of the cell cycle showed that PRMT5 knockdown caused G1 cell cycle arrest. (D) PRMT5 facilitated the G1 to S phase transition of the cell cycle in bladder cancer. Corresponding statistics of knockdown (E) and overexpression (F) are presented by a bar graph (*P < 0.05, **P < 0.01, ***P < 0.001). (G, H) Western blots were performed after knockdown or overexpression of PRMT5 to evaluate the relevant protein expression of the cell cycle and the mTOR signaling pathway.
Hypophosphorylated retinoblastoma (Rb) protein is a crucial regulator of cell transit from the G1 to S phase [18–20]. We investigated the phosphorylation status of Rb in the depletion and overexpression of PRMT5. The decreased level of phosphorylated Rb in BUC was consistent with the observation of PRMT5 depletion, suggesting that PRMT5 affects the cell cycle in BUC by controlling Rb activation (Figure 6G). p27 inhibits cyclin E activity, which causes hyperphosphorylation of Rb proteins [21–23]. Therefore, we examined whether PRMT5 depletion increased the expression of p27 in BUC via western blot analysis (Figure 6G).
Subsequently, we tested the impact of PRMT5 on the PI3K/Akt/mTOR pathway in BUC cells. Western blot analysis demonstrated that PRMT5 knockdown led to downregulation of PI3K, which deactivated Akt and mTOR phosphorylation. Conversely, PRMT5 upregulation promoted PI3K, resulting in activation of Akt and mTOR phosphorylation (Figure 6H)
Discussion
BUC is the seventh most common cancer in men worldwide. The TNM staging system is based on the extent of disease at presentation and clinicopathology and provides clinical prognostic indicators for BUC. However, patients with the same TNM stage and undergoing the same treatment display considerable variability in recurrence and survival [24]. Many BUC patients experience tumor recurrence and metastasis. Determination of a patient’s prognosis using molecular biomarkers before treatment may allow more effective use of adjuvant therapy, thus improving patients’ quality of life [25, 26]. Many novel oncogenes, such as chromosome 14 open reading frame 166 [27], trimethylation of lysine 27 on histone H3 [28], and maelstrom [5], promote malignant phenotypes in BUC. However, they are not precise enough to predict the patient’s prognosis.
Many arginine and lysine methyltransferases have been reported in cancers. Arginine methylation is an important regulator of biological function, tumorigenesis, and tumor progression [11, 14]. PRMT5 is the predominant type II methyltransferase and has a variety of functions. Together, these functions may contribute to increased tumorigenesis, leading to malignancies such as prostate, hepatocellular, and breast cancer and melanoma [12, 14]. PRMT5 and its substrate-binding partner WDR77 regulate alternative splicing through methylation of ZNF326 in breast cancer [17]. Upregulation of PRMT5 in glioblastoma cells increases their self-renewal capacity and proliferation through the PRMT5–PTEN molecular pathway [29]. These findings indicate that high PRMT5 expression promotes the proliferative and migratory processes of several types of solid cancer in humans. However, the role of PRMT5 in BUC oncogenesis had not been previously clarified. Thus, in this study, we explored the potential oncogenic role of PRMT5 in BUC. First, we observed that PRMT5 was significantly upregulated in BUC tissue compared with adjacent nontumor tissue. Recent reports have indicated that overexpression of the PRMT5 protein is positively correlated with advanced disease stage and adverse prognosis of certain human solid tumors [30, 31]. In patients with BUC, we also found that elevated PRMT5 was correlated with poor disease prognosis. As shown by Kaplan-Meier analysis and multivariate Cox analysis, PRMT5 was an independent adverse prognostic factor for overall and disease-free survival in patients with BUC. These findings indicate that PRMT5 may promote the growth and progression of BUC.
PRMT5 is a ubiquitous and evolutionarily conserved protein, but it may be involved in proliferation, migration, and invasion of cancer. PRMT5 upregulation results in increased proliferation and anchorage-independent colony growth, whereas cellular proliferation and colony formation in cancer are significantly inhibited by PRMT5 knockdown [14, 32–34]. Studies have considered direct inhibition resulting from increased translation [34, 35]. These reports indicated that there was one mechanism by which PRMT5 controls cell growth. Based on the upregulation of PRMT5 in BUC in our tests, we conducted phenotypic experiments and explored the potential biological role of PRMT5 in vivo and in vitro. In our experiments, PRMT5 knockdown resulted in significant inhibition of growth, migration, and invasion of BUC cells. Clearly, further study is needed to more accurately detect the signaling pathway involved in the oncogenic process of BUC induced by the upregulation of PRMT5. We conducted bioinformatics analysis, and the array showed that expression of PRMT5 in BUC was significantly associated with the cell cycle and the mTOR signaling pathway.
Distorted activity of proteins that affect the cell cycle induces abnormal proliferation, resulting in malignant transformation. We demonstrated that PRMT5 facilitated the progression of cells through the G1 to S phase, in accordance with the bioinformatics analysis. PRMT5 mediated the repression of p27 and the activation of Rb in BUC. Furthermore, our data showed that PRMT5 boosted the activity of the mTOR signaling pathway in BUC through upregulation of PI3K and Akt. Specifically, PRMT5 inhibition markedly impaired the PI3K/Akt/ mTOR pathway. PRMT5 may affect BUC tumorigenesis through this pathway. However, the relationship between PRMT5 and the PI3K/Akt/mTOR signaling pathway needs further research.
In summary, we demonstrated that PRMT5 upregulation in BUC is correlated with a poor prognosis and BUC proliferation, migration, and invasion. In addition, PRMT5 affects BUC tumor progression by distorting the Rb and p27 proteins involved in the cell cycle and facilitating the PI3K/Akt/mTOR signaling pathway. PRMT5 inhibitors may be a potential therapeutic approach that offers promising perspectives of BUC in the future.
Materials and Methods
Cell lines and cell culture
J82, UMUC, RT4, Biu87, TCCsup, 5637, and T24 bladder cancer cells from the American Type Culture Collection were cultured in RPMI 1640 medium or DMEM (Invitrogen, Carlsbad, CA, USA) with 10% fetal bovine serum (South Logan, UT, USA). All of cells were cultivated in an atmosphere at 37°C with 5% CO2.
qRT-PCR
RNA isolated with Trizol (Invitrogen) from the samples was quantified using an ultramicrospectrophotometer (NanoDrop ND-1000, USA), and it was reverse transcribed into cDNA using PrimeScriptTM Master Mix (TaKaRa, Japan). Then qRT-PCR was run on a Light-Cycler 480 instrument (Roche Diagnostics, Germany). The relative quantitative value was calculated using the 2-ΔΔCt method. The final result was recorded as fold change. Every sample was performed with three replicates.
Western blot and immunohistochemistry
All protein samples were extracted from the clinical bladder tissues and BUC cell lines using RIPA buffer (P00013C, Beyotime) and proteinase inhibitor cocktail (Roche, USA). The concentration of the protein was quantified using the BCA protein assay (Thermo Fisher Scientific, USA). Western blots were implemented in accordance with the standard methods [36]. Immunohistochemistry, which was used to determine the expression of PRMT5 in BUC tissues, has been described in previous studies [27]. An anti-PRMT5 antibody (1:500; Proteintech Group) was used.
RNA interference
To knock down the expression of PRMT5, two PRMT5 small-interfering RNAs (siRNAs) and the cognate control siRNA were synthesized by Gene Pharma (Shanghai, China). Then, 20 μm of siRNA was transfected into the indicated cells in six-well plates through the Lipofectamine 2000 Reagent (Invitrogen) according to the manufacturer’s instructions. The target sequences of PRMT5 for constructing the siRNAs are as follows: siRNA1, 5′-CCAGUUUGAGAUGCCUUAUT TAUAAGGCAUCUCAAACUGGGC-3′; siRNA2, 5′-GUUUCAAGAGGGAGUUCAUTTAUGAACUCCCUCUUGAAACGC-3′. Western blot was used to verify the effect of gene silencing 48 hours after transfection.
Cell counting kit-8 (CCK-8) assay
The indicated cells were sowed in each well at a density of 2,000 cells on 96-well plates. We examined the cell growth every 24 hours with CCK-8 (Beyotime Technology, China) after incubation for 30 minutes at 37°C. According to the reference wavelength, the absorbance value was detected at 450 nm.
Colony formation assay
The indicated cells were sowed in each well at a density of 500 cells on six-well plates. After cultivation for 7 days, the colonies were stained with crystal violet and visualized with a microscope.
5-Ethynyl-2'-deoxyuridine (EdU) assay
5-Ethynyl-20-deoxyuridine (EdU), a more specific and sensitive method, was mainly used as the marker of cellular replication activity [37]. For an EdU incorporation assay, cells were placed on every confocal well (2,000 cells per well) and cultured for 24 hours. EdU fluorescent staining was performed using a commercial cell-light EdU Apollo 567 in vitro kit (RIB&BIO, Guangzhou, China) according to the manufacturer’s protocol. A fluorescence confocal microscope (Nikon, Japan) was used to directly detect staining for PRMT5 in the EdU-labeled cells.
Transwell invasion assay
Biu87 and T24 cells (5×104 cells/well) were plated on the upper chamber of 24-well plates in RPMI 1640 without serum. Then 20% formaldehyde was used to fix the invading cells, and crystal violet was used for staining at 24 hours after seeding.
Cell wound-healing assay
The cell monolayer in six-well plates was scraped by a sterile 100-mL pipette tip and then washed with PBS. The scratched wells were cultivated in a serum-free medium. At the time points of 0 and 24 hours, the scratched cell images were captured to analyze migration by an inverted microscope.
Tumor growth in xenografts
Twelve male BALB/c nude mice were divided into two groups. They were fed with sterilized feed and pure water in a specified pathogen-free environment with air laminar flow chamber. Feed, bedding, and cages for indoor use were autoclaved and transported through a sterile inlet chamber to exclude any microorganisms. Then mice were divided into two groups and inoculated subcutaneously with 1×107 of T24-shRNA or T24-negative control cells. The xenografts were observed and sized using Vernier calipers every week after injection. Five weeks later, all of the mice were euthanized by cervical dislocation, and the subcutaneous tumor xenografts in the mice were collected. The weight and maximum diameter of the tumor xenografts were measured using an electronic scale and Vernier calipers, respectively. Finally, the tumor xenografts were fixed in 10% formalin and embedded in paraffin for hematoxylin and eosin staining and the histopathological analysis.
Cell cycle analysis
Double thymidine was used for synchronization. Posttreatment cells were treated with the cell cycle analysis kit (4Abio, Beijing, China) according to the standard protocol. Stained cells were tested using a flow cytometer (cytoFLEX, Beckman, USA).
Patient and specimen characteristics
PRMT5 expression data and detailed clinical information of BUC patients were first ascertained from the data sets of the Oncomine database (https://www.oncomine.org) and The Cancer Genome Atlas (https://www.cancer.gov/about-nci/organization/ccg/research/structural-genomics/tcga). One hundred thirty-two clinical BUC samples from Chinese patients from 2002 to 2016 were collected at the Sun Yat-Sen University Cancer Center. The clinical and pathological information collected included age, gender, surgery, tumor size, TNM stage, differentiation grade, vital status, and follow-up time, and tumors were classified according to the guidelines of the European Association of Urology [39]. All samples were from patients who had not yet received chemotherapy, radiotherapy, or immunotherapy and were obtained after surgery. All samples were acquired after receiving informed consent and approval from the Ethical Committee of Sun Yat-Sen University Cancer Center (Guangzhou, China).
Statistical analysis
Using SPSS 20.0 (SPSS, Inc., Chicago, IL, USA) and GraphPad Prism 6.0 (GraphPad Software, La Jolla, CA, USA), the mRNA levels of PRMT5 in BUC tumor tissues were compared with those in adjacent nontumor tissues using a paired student’s t test. Relationships between PRMT5 expression and clinical factors were analyzed. Descriptive statistics and the χ2 test were used to assess the differences between groups. Overall survival was analyzed by the Kaplan-Meier method using the log-rank test. Univariate and multivariate Cox regression analyses were performed to assess the correlation among age, gender, tumor size, tumor stage, and differentiation grade. P values < 0.05 were considered statistically significant.
Ethics approval
The Ethical Committee of Sun Yat-Sen University Cancer Center provided permission to use the clinical samples. All patients provided written consent for the clinical samples and data to be used for scientific research. The animal experiment was approved by the Animal Ethical and Welfare Committee of Sun Yat-Sen University and abided by the Guide for the Care and Use of Laboratory Animals.
Author Contributions
JL and ZQ designed and conceived the experiments. LT, KX, YY, and HL performed the experiments. LT, KX, YY, MC, ZQ, and JL collected patient specimens and account for the data. LT, KX, YY, JL, and ZQ wrote the paper. LT, KX, and YY contributed equally to the work. All authors read and approved the final manuscript.
Acknowledgments
We thank LetPub (www.letpub.com) for its linguistic assistance during the preparation of this manuscript.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Funding
This study was supported by grants from the Natural Science Foundation of Guangdong Province of China (Grant No. 2016A030310213), mainly used to conceive and determine the experiments; the National Natural Science Foundation of China (Grant No. 81202013); and the Guangdong Science and Technology Department Project (Grant No. 2017A020215150), mainly used for collection of the samples, analysis, and interpretation of data. All of the funds were applied to labor costs for the work performed in the study and the writing of the manuscript.
References
- 1. Li K, Lin T, Xue W, Mu X, Xu E, Yang X, Chen F, Li G, Ma L, Wang G, Liang C, Shi H, Li M, et al, and Chinese Bladder Cancer Consortium. Current status of diagnosis and treatment of bladder cancer in China - Analyses of Chinese Bladder Cancer Consortium database. Asian J Urol. 2015; 2:63–69. https://doi.org/10.1016/j.ajur.2015.04.016 [PubMed]
- 2. Antoni S, Ferlay J, Soerjomataram I, Znaor A, Jemal A, Bray F. Bladder Cancer Incidence and Mortality: A Global Overview and Recent Trends. Eur Urol. 2017; 71:96–108. https://doi.org/10.1016/j.eururo.2016.06.010 [PubMed]
- 3. Alfred Witjes J, Lebret T, Compérat EM, Cowan NC, De Santis M, Bruins HM, Hernández V, Espinós EL, Dunn J, Rouanne M, Neuzillet Y, Veskimäe E, van der Heijden AG, et al. Updated 2016 EAU Guidelines on Muscle-invasive and Metastatic Bladder Cancer. Eur Urol. 2017; 71:462–75. https://doi.org/10.1016/j.eururo.2016.06.020 [PubMed]
- 4. Knowles MA, Hurst CD. Molecular biology of bladder cancer: new insights into pathogenesis and clinical diversity. Nat Rev Cancer. 2015; 15:25–41. https://doi.org/10.1038/nrc3817 [PubMed]
- 5. Li XD, Zhang JX, Jiang LJ, Wang FW, Liu LL, Liao YJ, Jin XH, Chen WH, Chen X, Guo SJ, Zhou FJ, Zeng YX, Guan XY, et al. Overexpression of maelstrom promotes bladder urothelial carcinoma cell aggressiveness by epigenetically downregulating MTSS1 through DNMT3B. Oncogene. 2016; 35:6281–92. https://doi.org/10.1038/onc.2016.165 [PubMed]
- 6. Li Y, Liu H, Lai C, Du X, Su Z, Gao S. The Lin28/let-7a/c-Myc pathway plays a role in non-muscle invasive bladder cancer. Cell Tissue Res. 2013; 354:533–41. https://doi.org/10.1007/s00441-013-1715-6 [PubMed]
- 7. Pollack A, Wu CS, Czerniak B, Zagars GK, Benedict WF, McDonnell TJ. Abnormal bcl-2 and pRb expression are independent correlates of radiation response in muscle-invasive bladder cancer. Clin Cancer Res. 1997; 3:1823–29. [PubMed]
- 8. van Rhijn BW, Lurkin I, Radvanyi F, Kirkels WJ, van der Kwast TH, Zwarthoff EC. The fibroblast growth factor receptor 3 (FGFR3) mutation is a strong indicator of superficial bladder cancer with low recurrence rate. Cancer Res. 2001; 61:1265–68. [PubMed]
- 9. Sakr SA, Mahran HA, Fahmy AM, El-Kholy MA, Meawad M. Expression of c-erb-B2 gene in bladder cancer of Egyptian patients and its correlation with p53 and bcl-2. Biomed Pharmacother. 2015; 76:73–81. https://doi.org/10.1016/j.biopha.2015.10.021 [PubMed]
- 10. Xie R, Chen X, Chen Z, Huang M, Dong W, Gu P, Zhang J, Zhou Q, Dong W, Han J, Wang X, Li H, Huang J, Lin T. Polypyrimidine tract binding protein 1 promotes lymphatic metastasis and proliferation of bladder cancer via alternative splicing of MEIS2 and PKM. Cancer Lett. 2019; 449:31–44. https://doi.org/10.1016/j.canlet.2019.01.041 [PubMed]
- 11. Blanc RS, Richard S. Arginine Methylation: The Coming of Age. Mol Cell. 2017; 65:8–24. https://doi.org/10.1016/j.molcel.2016.11.003 [PubMed]
- 12. Kryukov GV, Wilson FH, Ruth JR, Paulk J, Tsherniak A, Marlow SE, Vazquez F, Weir BA, Fitzgerald ME, Tanaka M, Bielski CM, Scott JM, Dennis C, et al. MTAP deletion confers enhanced dependency on the PRMT5 arginine methyltransferase in cancer cells. Science. 2016; 351:1214–18. https://doi.org/10.1126/science.aad5214 [PubMed]
- 13. Sheng X, Wang Z. Protein arginine methyltransferase 5 regulates multiple signaling pathways to promote lung cancer cell proliferation. BMC Cancer. 2016; 16:567. https://doi.org/10.1186/s12885-016-2632-3 [PubMed]
- 14. Stopa N, Krebs JE, Shechter D. The PRMT5 arginine methyltransferase: many roles in development, cancer and beyond. Cell Mol Life Sci. 2015; 72:2041–59. https://doi.org/10.1007/s00018-015-1847-9 [PubMed]
- 15. Kanda M, Shimizu D, Fujii T, Tanaka H, Shibata M, Iwata N, Hayashi M, Kobayashi D, Tanaka C, Yamada S, Nakayama G, Sugimoto H, Koike M, et al. Protein arginine methyltransferase 5 is associated with malignant phenotype and peritoneal metastasis in gastric cancer. Int J Oncol. 2016; 49:1195–202. https://doi.org/10.3892/ijo.2016.3584 [PubMed]
- 16. Deng X, Shao G, Zhang HT, Li C, Zhang D, Cheng L, Elzey BD, Pili R, Ratliff TL, Huang J, Hu CD. Protein arginine methyltransferase 5 functions as an epigenetic activator of the androgen receptor to promote prostate cancer cell growth. Oncogene. 2017; 36:1223–31. https://doi.org/10.1038/onc.2016.287 [PubMed]
- 17. Rengasamy M, Zhang F, Vashisht A, Song WM, Aguilo F, Sun Y, Li S, Zhang W, Zhang B, Wohlschlegel JA, Walsh MJ. The PRMT5/WDR77 complex regulates alternative splicing through ZNF326 in breast cancer. Nucleic Acids Res. 2017; 45:11106–20. https://doi.org/10.1093/nar/gkx727 [PubMed]
- 18. Bartek J, Bartkova J, Lukas J. The retinoblastoma protein pathway in cell cycle control and cancer. Exp Cell Res. 1997; 237:1–6. https://doi.org/10.1006/excr.1997.3776 [PubMed]
- 19. Qian Y, Luckey C, Horton L, Esser M, Templeton DJ. Biological function of the retinoblastoma protein requires distinct domains for hyperphosphorylation and transcription factor binding. Mol Cell Biol. 1992; 12:5363–72. https://doi.org/10.1128/MCB.12.12.5363 [PubMed]
- 20. Vandel L, Nicolas E, Vaute O, Ferreira R, Ait-Si-Ali S, Trouche D. Transcriptional repression by the retinoblastoma protein through the recruitment of a histone methyltransferase. Mol Cell Biol. 2001; 21:6484–94. https://doi.org/10.1128/MCB.21.19.6484-6494.2001 [PubMed]
- 21. Kato J, Matsushime H, Hiebert SW, Ewen ME, Sherr CJ. Direct binding of cyclin D to the retinoblastoma gene product (pRb) and pRb phosphorylation by the cyclin D-dependent kinase CDK4. Genes Dev. 1993; 7:331–42. https://doi.org/10.1101/gad.7.3.331 [PubMed]
- 22. Hinds PW, Mittnacht S, Dulic V, Arnold A, Reed SI, Weinberg RA. Regulation of retinoblastoma protein functions by ectopic expression of human cyclins. Cell. 1992; 70:993–1006. https://doi.org/10.1016/0092-8674(92)90249-C [PubMed]
- 23. Sherr CJ, Roberts JM. CDK inhibitors: positive and negative regulators of G1-phase progression. Genes Dev. 1999; 13:1501–12. https://doi.org/10.1101/gad.13.12.1501 [PubMed]
- 24. Schrier BP, Hollander MP, van Rhijn BW, Kiemeney LA, Witjes JA. Prognosis of muscle-invasive bladder cancer: difference between primary and progressive tumours and implications for therapy. Eur Urol. 2004; 45:292–96. https://doi.org/10.1016/j.eururo.2003.10.006 [PubMed]
- 25. Hussain SA, James ND. Molecular markers in bladder cancer. Semin Radiat Oncol. 2005; 15:3–9. https://doi.org/10.1016/j.semradonc.2004.07.007 [PubMed]
- 26. Wang C, Liu Q, Huang M, Zhou Q, Zhang X, Zhang J, Xie R, Yu Y, Chen S, Fan J, Chen X. Loss of GATA6 expression promotes lymphatic metastasis in bladder cancer. FASEB J. 2020; 34:5754–66. https://doi.org/10.1096/fj.201903176R [PubMed]
- 27. Chen M, Ye Y, Zou B, Guo S, Zhou F, Lu K, Liu J, Xu Z, Han H, Liu Z, Li Y, Yao K, Liu C, Qin Z. C14orf166 is a high-risk biomarker for bladder cancer and promotes bladder cancer cell proliferation. J Transl Med. 2016; 14:55. https://doi.org/10.1186/s12967-016-0801-4 [PubMed]
- 28. Liu J, Li Y, Liao Y, Mai S, Zhang Z, Liu Z, Jiang L, Zeng Y, Zhou F, Xie D. High expression of H3K27me3 is an independent predictor of worse outcome in patients with urothelial carcinoma of bladder treated with radical cystectomy. Biomed Res Int. 2013; 2013:390482. https://doi.org/10.1155/2013/390482 [PubMed]
- 29. Banasavadi-Siddegowda YK, Russell L, Frair E, Karkhanis VA, Relation T, Yoo JY, Zhang J, Sif S, Imitola J, Baiocchi R, Kaur B. PRMT5-PTEN molecular pathway regulates senescence and self-renewal of primary glioblastoma neurosphere cells. Oncogene. 2017; 36:263–74. https://doi.org/10.1038/onc.2016.199 [PubMed]
- 30. Shimizu D, Kanda M, Sugimoto H, Shibata M, Tanaka H, Takami H, Iwata N, Hayashi M, Tanaka C, Kobayashi D, Yamada S, Nakayama G, Koike M, et al. The protein arginine methyltransferase 5 promotes malignant phenotype of hepatocellular carcinoma cells and is associated with adverse patient outcomes after curative hepatectomy. Int J Oncol. 2017; 50:381–86. https://doi.org/10.3892/ijo.2017.3833 [PubMed]
- 31. Wu Y, Wang Z, Zhang J, Ling R. Elevated expression of protein arginine methyltransferase 5 predicts the poor prognosis of breast cancer. Tumour Biol. 2017; 39:1010428317695917. https://doi.org/10.1177/1010428317695917 [PubMed]
- 32. Scoumanne A, Zhang J, Chen X. PRMT5 is required for cell-cycle progression and p53 tumor suppressor function. Nucleic Acids Res. 2009; 37:4965–76. https://doi.org/10.1093/nar/gkp516 [PubMed]
- 33. Wei TY, Juan CC, Hisa JY, Su LJ, Lee YC, Chou HY, Chen JM, Wu YC, Chiu SC, Hsu CP, Liu KL, Yu CT. Protein arginine methyltransferase 5 is a potential oncoprotein that upregulates G1 cyclins/cyclin-dependent kinases and the phosphoinositide 3-kinase/AKT signaling cascade. Cancer Sci. 2012; 103:1640–50. https://doi.org/10.1111/j.1349-7006.2012.02367.x [PubMed]
- 34. Pal S, Vishwanath SN, Erdjument-Bromage H, Tempst P, Sif S. Human SWI/SNF-associated PRMT5 methylates histone H3 arginine 8 and negatively regulates expression of ST7 and NM23 tumor suppressor genes. Mol Cell Biol. 2004; 24:9630–45. https://doi.org/10.1128/MCB.24.21.9630-9645.2004 [PubMed]
- 35. Karkhanis V, Hu YJ, Baiocchi RA, Imbalzano AN, Sif S. Versatility of PRMT5-induced methylation in growth control and development. Trends Biochem Sci. 2011; 36:633–41. https://doi.org/10.1016/j.tibs.2011.09.001 [PubMed]
- 36. Chen X, Xie R, Gu P, Huang M, Han J, Dong W, Xie W, Wang B, He W, Zhong G, Chen Z, Huang J, Lin T. Long Noncoding RNA LBCS Inhibits Self-Renewal and Chemoresistance of Bladder Cancer Stem Cells through Epigenetic Silencing of SOX2. Clin Cancer Res. 2019; 25:1389–403. https://doi.org/10.1158/1078-0432.CCR-18-1656 [PubMed]
- 37. Ligasová A, Liboska R, Friedecký D, Mičová K, Adam T, Oždian T, Rosenberg I, Koberna K. Dr Jekyll and Mr Hyde: a strange case of 5-ethynyl-2′-deoxyuridine and 5-ethynyl-2′-deoxycytidine. Open Biol. 2016; 6:150172. https://doi.org/10.1098/rsob.150172 [PubMed]
- 38. Wang LL, Jin XH, Cai MY, Li HG, Chen JW, Wang FW, Wang CY, Hu WW, Liu F, Xie D. AGBL2 promotes cancer cell growth through IRGM-regulated autophagy and enhanced Aurora A activity in hepatocellular carcinoma. Cancer Lett. 2018; 414:71–80. https://doi.org/10.1016/j.canlet.2017.11.003 [PubMed]
- 39. Babjuk M, Böhle A, Burger M, Capoun O, Cohen D, Compérat EM, Hernández V, Kaasinen E, Palou J, Rouprêt M, van Rhijn BW, Shariat SF, Soukup V, et al. EAU Guidelines on Non-Muscle-invasive Urothelial Carcinoma of the Bladder: Update 2016. Eur Urol. 2017; 71:447–61. https://doi.org/10.1016/j.eururo.2016.05.041 [PubMed]