



Introduction
Infertility has become a worldwide problem, affecting up to 20% of couples trying to conceive [1, 2]. In this context, a few important facts should be emphasized: 1) male factors (coexisting with female factors) contribute to infertility in up to 20–70% of cases, and one-third of these cases are due to male factors alone [1–3]; 2) an actual decline in semen quality over the past decades has been observed globally [4]; and 3) paternal age is rising, as an increasing number of men are decide to became a father at an older age [5, 6].
It is known that the risk of poor reproductive outcomes can increase with a male age of >40 or even >35 y, commonly classified as advanced age. Age-dependent changes in male organism (e.g. vascular sufficiency, increase in incidence of systemic diseases and infections, disorders of histological structure of testes, decreased levels of sex hormones, oxidative stress, de novo mutations) are deleterious and the consequences of advanced paternal age include a prolonged waiting time to pregnancy, delayed embryo development in in vitro conditions, an increased incidence of embryo implantation failure and abortions, pregnancy problems and live birth outcome [7–9] (Figure 1). In addition, advanced paternal age also seems to affect children's health. There is a positive correlation between paternal age and the incidence of mental deprivation of offspring, such as those associated with the autism spectrum and diseases such as schizophrenia, especially when the paternal age is ≥40–50 y [10–13]. Furthermore, the frequency of genetic disorders, such as Klinefelter syndrome [14]; Down syndrome, when mother age is >35 y [15]; and autosomal dominant diseases such as Marfan syndrome (men >40 y), Pfeiffer and Crouzon syndrome (men >50 y), Apert syndrome (men >37 y), achondroplasia and neurofibromatosis type 1 rises in children of fathers >40 y. Unfortunately, the risk of central nervous system and breast cancers as well as leukaemia is growing [12]. More often, children with heart defects (e.g., ventricular septal defects, atrial septal defects, large vessel transposition), neural tube defects, anencephaly and tracheo-oesophageal fistula have been born to men >35, 40, and 45 y of age [13] (Figure 1).
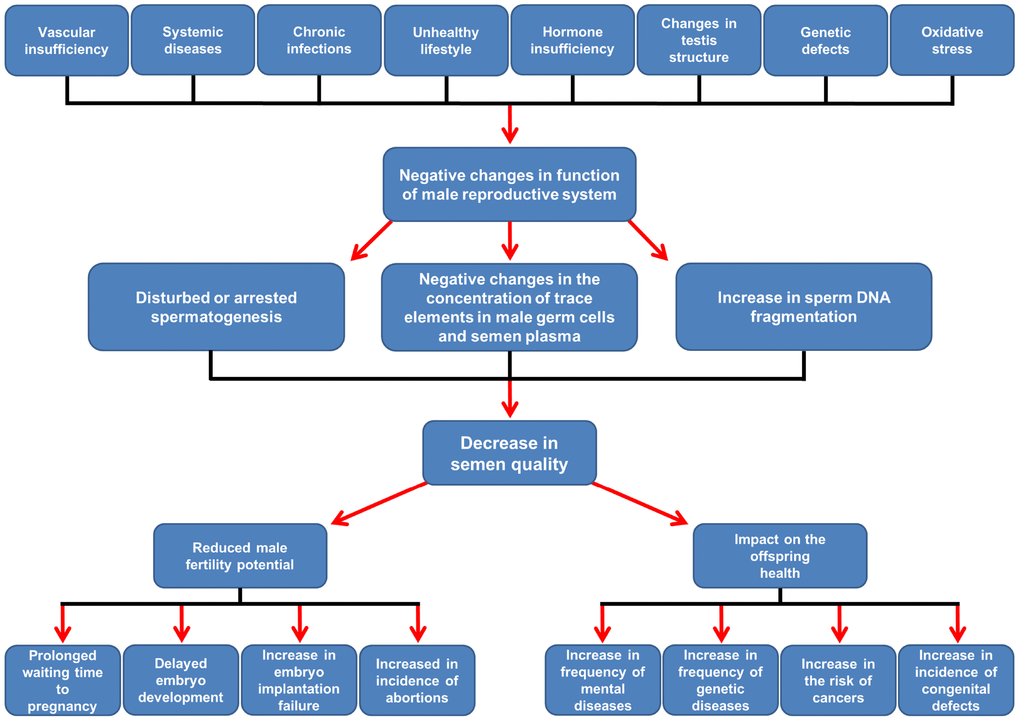
Figure 1. Scheme of suggested age-related changes affecting the male reproductive system and their consequences for male fertility and offspring health. Details in the text. According to Rosiak et al. [6, 9], modified.
Age-related negative changes in sperm quality are observed in men over 35 y of age, and with age (>40 y), these changes become more pronounced [6]. Deterioration of semen quality is visible in the ejaculate volume, sperm count, motility, vitality and sperm morphology [16, 17]. It should be highlighted that the detrimental effect of ageing is also noted in the sperm chromatin status [18, 19]. However, negative changes in all semen parameters are not always observed at the same time. Therefore, it is difficult to definitively determine the influence of age on male gonad function and semen quality. Due to unclear, ambiguous and controversial data, our study was designed to estimate the impact of paternal age not only on conventional semen characteristics but also on sperm nuclear DNA integrity.
Results
According to other authors [20–24] from the entire cohort of men and in the groups of men with normozoospermia and abnormal semen parameters, we decided to designate two subgroups of men: ≥40 y and <40 y of age. The obtained data revealed the differences in selected sperm conventional characteristics and sperm DNA fragmentation between age groups. The older men had a significantly lower ejaculate volume, percentage of sperm cells with normal morphology, and percentage of male gametes with non-progressive motility and a higher percentage of DFI (sperm DNA fragmentation index) than the younger men (Table 1). In turn, in the older subgroup with normozoospermia, the ejaculate volume and TZI (teratozoospermia index) were significantly decreased, but the percentage of spermatozoa with progressive motility, percentage of eosin-negative spermatozoa and percentage of male gametes with DNA strand breaks were significantly increased compared to those of the younger subgroup with normozoospermia (Table 2). Moreover, the older subgroup with abnormal semen parameters had a significantly lower ejaculate volume and percentage of sperm cells with non-progressive motility and a higher percentage of DFI than the younger subgroup with abnormal semen characteristics (Table 3).
Table 1. Descriptive statistics and comparisons of standard semen parameters and DFI between men aged <40 y and ≥40 y.
| Parameters n median (range) mean±SD | < 40 y (n = 902) | ≥ 40 y (n = 222) | p | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Age (y) | 902 32.00 (18.00–39.00) 31.05 (5.07) | 222 43 (40.00–70.00) 45.41 (6.11) | 0.00001 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Semen volume (mL) | 902 3.35 (0.50–12.00) 3.54 (1.66) | 222 3.00 (0.15–10.00) 3.10 (1.63) | 0.00012 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Sperm concentration (×106/mL) | 902 33.56 (0.001–347.700) 47.03 (44.46) | 222 39.50 (0.10–216.00) 50.97 (46.57) | 0.40039 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Total number of spermatozooa (x106) | 902 107.70 (0.002–2109.75) 166.22 (191.67) | 222 105.50 (0.15–931.50) 148.72 (154.44) | 0.2374 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Morphologically normal spermatozoa (%) | 902 4.00 (0.00–34.00) 5.63 (5.80) | 222 4.00 (0.00–28.00) 4.36 (4.14) | 0.03200 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TZI | 845 1.63 (1.02–2.62) 1.65 (0.25) | 218 1.61 (1.21–2.33) 1.62 (0.21) | 0.11902 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Progressive motility (%) | 902 40.00 (0.00–90.00) 39.32 (19.73) | 222 41.00 (0.00–88.00) 40.62 (21.76) | 0.40505 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Non-progressive motility (%) | 902 14.00 (0.00–59.00) 16.38 (9.80) | 222 11.00 (0.00–54.00) 13.71 (8.86) | 0.00013 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Immotile sperms (%) | 902 41.00 (2.00–100.00) 44.27 (19.15) | 222 44.00 (1.00–99.00) 45.66 (23.12) | 0.61195 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Eosin-negative spermatozoa – live cells (%) | 897 68.00 (0.00–98.00) 65.18 (17.73) | 220 66.00 (2.00–99.00) 64.41 (20.76) | 0.95651 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Hos-test-positive spermatozoa – live cells (%) | 631 62.00 (0.00–93.00) 59.25 (17.95) | 190 65.00 (10.00–94.00) 60.24 (18.19) | 0.47225 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Round cells concentration (x 106/mL) | 862 0.16 (0.00–37.34) 0.74 (2.16) | 216 0.10 (0.00–11.00) 0.51 (1.22) | 0.21214 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Peroxidase-positive cells (x106/mL) | 869 0.00 (0.00–11.70) 0.37 (1.06) | 216 0.00 (0.00–6.00) 0.28 (0.74) | 0.27913 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DFI (%) | 248 10.82 (0.77–65.14) 13.37 (10.31) | 88 18.47 (0.76–75.94) 19.77 (13.12) | 0.00001 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DFI – sperm DNA fragmentation index, n – number of subjects, SD – standard deviation, TZI – teratozoospermia index, p – significance of differences between compared groups, Mann–Whitney U test | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Table 2. Descriptive statistics and comparisons of standard semen parameters and DFI between men aged <40 y and ≥40 y with normozoospermia.
| Parameters n median (range) mean±SD | < 40 y (n = 367) | ≥ 40 y (n = 81) | p | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Age (y) | 367 31.00 (18.00–39.00) 30.06 (5.25) | 81 43.00 (40.00–53.00) 43.97 (4.10) | 0.00001 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Semen volume (mL) | 367 3.50 (0.80–12.00) 3.86 (1.77) | 81 2.80 (0.80–8.70) 3.30 (1.68) | 0.00219 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Sperm concentration (×106/mL) | 367 56.00 (8.90–347.70) 68.05 (50.58) | 81 59.00 (9.20–216.00) 72.49 (46.33) | 0.24677 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Total number of spermatozooa (x106) | 367 187.00 (39.00–2109.75) 251.49 (235.54) | 81 171.00 (39.00–931.50) 225.14 (170.72) | 0.56813 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Morphologically normal spermatozoa (%) | 367 7.00 (4.00–34.00) 9.40 (6.33) | 81 7.00 (4.00–28.00) 7.62 (4.31) | 0.10451 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TZI | 350 1.54 (1.08–2.49) 1.54 (0.20) | 79 1.49 (1.21–1.89) 1.49 (0.15) | 0.03818 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Progressive motility (%) | 367 51.00 (32.00–90.00) 52.41 (12.94) | 81 55.00 (32.00–88.00) 56.22 (13.67) | 0.02615 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Non-progressive motility (%) | 367 14.00 (1.00–44.00) 15.27 (8.03) | 81 11.00 (3.00–33.00) 13.58 (7.22) | 0.11685 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Immotile sperms (%) | 367 32.00 (2.00–66.00) 32.44 (11.63) | 81 30.00 (1.00–58.00) 30.19 (13.05) | 0.23924 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Eosin-negative spermatozoa – live cells (%) | 366 74.00 (35.00–98.00) 73.19 (11.34) | 81 80.00 (46.00–99.00) 76.37 (13.23) | 0.01771 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Hos-test-positive spermatozoa – live cells (%) | 252 69.50 (28.00–92.00) 67.40 (11.91) | 73 69.00 (40.00–94.00) 69.49 (11.87) | 0.22170 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Round cells concentration (x 106/mL) | 351 0.15 (0.00–37.34) 0.83 (2.88) | 79 0.10 (0.00–11.70) 0.59 (1.52) | 0.97768 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Peroxidase-positive cells (x106/mL) | 354 0.00 (0.00–11.56) 0.28 (0.86) | 79 0.00 (0.00–6.00) 0.25 (0.75) | 0.81045 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DFI (%) | 116 11.45 (0.77–65.14) 12.74 (9.48) | 44 14.77 (1.00–40.52) 17.68 (10.45) | 0.00518 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DFI – sperm DNA fragmentation index, n – number of subjects, SD – standard deviation, TZI – teratozoospermia index, p – significance of differences between compared groups, Mann–Whitney U test | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Table 3. Descriptive statistics and comparisons of standard semen parameters and DFI between men aged <40 y and ≥40 y with abnormal semen parameters.
| Parameters n median (range) mean±SD | < 40 y (n = 535) | ≥ 40 y (n = 141) | p | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Age (y) | 535 32.00 (18.00–39.00) 31.72 (4.83) | 141 44.00 (40.00–70.00) 46.24 (6.88) | 0.00001 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Semen volume (mL) | 535 3.00 (0.50–10.00) 3.32 (1.55) | 141 3.00 (0.15–10.00) 2.99 (1.60) | 0.01952 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Sperm concentration (×106/mL) | 535 22.50 (0.001–238.30) 32.62 (32.71) | 141 22.30 (0.10–210.00) 38.61 (42.14) | 0.55474 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Total number of spermatozooa (x106) | 535 62.50 (0.002–800.00) 107.74 (124.64) | 141 59.50 (0.15–689.00) 104.82 (125.21) | 0.44229 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Morphologically normal spermatozoa (%) | 535 2.00 (0.00–22.00) 3.04 (3.58) | 141 2.00 (0.00–11.00) 2.48 (2.61) | 0.24004 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TZI | 495 1.72 (1.02–2.62) 1.73 (0.24) | 139 1.68 (1.26–2.33) 1.70 (0.21) | 0.16066 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Progressive motility (%) | 535 27.00 (0.00–84.00) 30.34 (18.53) | 141 30.00 (0.00–88.00) 31.65 (20.47) | 0.59229 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Non-progressive motility (%) | 535 15.00 (0.00–59.00) 17.14 (10.79) | 141 11.00 (0.00–54.00) 13.79 (9.70) | 0.00033 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Immotile sperms (%) | 535 50.00 (6.00–100.00) 52.38 (19.09) | 141 52.00 (4.00–99.00) 54.55 (22.99) | 0.21761 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Eosin-negative spermatozoa – live cells (%) | 531 64.00 (0.00–98.00) 59.66 (19.19) | 139 58.00 (2.00–97.00) 57.44 (21.21) | 0.23521 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Hos-test-positive spermatozoa – live cells (%) | 379 57.00 (0.00–90.00) 53.84 (19.21) | 117 57.00 (11.00–90.00) 54.47 (19.09) | 0.84956 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Round cells concentration (x 106/mL) | 511 0.20 (0.00–14.68) 0.68 (1.48) | 137 0.10 (0.00–7.03) 0.46 (1.00) | 0.10633 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Peroxidase-positive cells (x106/mL) | 515 0.00 (0.00–11.70) 0.43 (1.17) | 137 0.00 (0.00–4.80) 0.30 (0.74) | 0.21906 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DFI (%) | 132 10.54 (1.00–60.20) 13.92 (10.99) | 44 20.45 (0.76–75.94) 21.85 (15.18) | 0.00120 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DFI – sperm DNA fragmentation index, n – number of subjects, SD – standard deviation, TZI – teratozoospermia index, p – significance of differences between compared groups, Mann–Whitney U test | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Furthermore, based on reports by other authors [25–27], we decided to designate a cut-off point for DNA damage of 10%. This threshold is considered to indicate high sperm DNA damage and is referred to as low fertility potential. The older men in the entire cohort, the group of men with normozoospermia and the group of men with abnormal semen parameters had a significantly higher prevalence of sperm cells with DNA damage >10% than the younger men in these groups (subjects: 72.73% vs. 53.63%, 75.00% vs. 55.17%, 70.46% vs. 52.27%, respectively) (Tables 4–6). Likewise, men ≥40 y of age had an OR (odds ratio) for having a high level of sperm DNA damage that was over 2-fold higher than that of the men <40 y in both the entire cohort and the group of men with normal and abnormal standard semen parameters (OR: 2.3058, OR: 2.4375, OR: 2.1773, respectively) (Tables 7–9).
Table 4. Prevalence of DFI in the group of men aged ≥40 y and <40 y in the entire cohort.
| Group | DFI (%) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0–10% n(%) | >10% n(%) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| <40 y (n = 248) | 115(46.37) | 133(53.63) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ≥40 y (n = 88) | 24(27.27)* | 64(72.73)* | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DFI – sperm DNA fragmentation index, n – number of subjects. *Significant difference between compared groups at p = 0.0018; chi-squared test | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Table 5. Prevalence of DFI in the group of men aged ≥40 y and <40 y with normal standard semen parameters.
| Group | DFI (%) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0–10% n(%) | >10% n(%) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| <40 y (n = 116) | 52(44.83) | 64(55.17) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ≥40 y (n = 44) | 11(25.00)* | 33(75.00)* | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DFI – sperm DNA fragmentation index, n – number of subjects. *Significant difference between compared groups at p = 0.0223; chi-squared test | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Table 6. Prevalence of DFI in the group of men aged ≥40 y and <40 y with abnormal standard semen parameters.
| Group | DFI (%) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0–10% n(%) | >10% n(%) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| <40 y (n = 132) | 63(47.73) | 69(52.27) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ≥40 y (n = 44) | 13(29.54)* | 31(70.46)* | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DFI – sperm DNA fragmentation index, n – number of subjects. *Significant difference between compared groups at p = 0.0355; chi-squared test. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Table 7. Odds ratio (OR) for DFI in the group of men aged ≥40 y compared to men <40 y in the entire cohort.
| < 40 y (n = 248) n(%) | ≥ 40 y (n = 88) n(%) | OR (95%CI) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DFI 0–10% | 115(46.37) | 24(27.27) | 0.4337* (0.2549 to 0.7378) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DFI >10% | 133(53.63) | 64(72.73) | 2.3058* (1.3553–3.9228) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DFI – sperm DNA fragmentation index, n – number of subjects. *Statistical significance at p = 0.0021; 95% CI – 95% confidential interval | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Table 8. Odds ratio (OR) for DFI in the group of men aged ≥ 40 y compared to men <40 y with normal standard semen parameters.
| <40 (n = 116) n(%) | ≥40 y (n = 44) (n%) | OR (95%CI) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DFI 0–10% | 52(44.83) | 11(25.00) | 0.4103* (0.1891–0.8899) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DFI >10% | 64(55.17) | 33(75.00) | 2.4375* (1.1237–5.2871) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DFI – sperm DNA fragmentation index, n – number of subjects.*statistical significance at p = 0.0241; 95% CI – 95% confidential interval | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Table 9. Odds ratio (OR) for DFI in the group of men aged ≥40 y compared to men <40 y with abnormal standard semen parameters.
| <40 y (n = 132) n(%) | ≥40 y (n = 44) n(%) | OR (95%CI) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DFI 0–10% | 63(47.73) | 13(29.54) | 0.4593* (0.2209–0.9551) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DFI >10% | 69(52.27) | 31(70.46) | 2.1773* (1.0470–4.5278) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DFI – sperm DNA fragmentation index, n – number of subjects. *statistical significance at p = 0.0373; 95% CI – 95% confidential interval | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Discussion
Age-dependent decrease in selected semen characteristics
In our study, we wanted to verify the influence of ageing on semen quality. Therefore, based on the suggestions of other authors [20, 22, 23] men ≥40 y and <40 y of age were distinguished in the entire cohort of subjects, in the group of men with normozoospermia and in the group of men with abnormal semen parameters. We observed a significant decrease in semen volume with ageing. This parameter declined after ≥40 y of age in all study groups. The obtained results were in line with those of other researchers [16, 28]. However, an association between age and semen volume was not always found [7, 20, 23].
In accordance with most of the published reports [7, 16], we found that ageing is characterized by a statistically significant decrease in the percentages of morphologically normal spermatozoa (in the entire cohort). Stone et al. [16] demonstrated a decline in the percentage of sperm cells with normal morphology in men aged >40 y. Similar results were obtained by other authors [7, 29, 30] who have shown that the percentage of sperm cells with normal structure decreased significantly in men aged >50–79 y. However, Park et al. [31] and Kaarouch et al. [20] did not show a relationship between sperm morphology and paternal age.
In the present study, no significant age-related decreases in the sperm concentration, total sperm count, sperm motility or vitality were revealed, which was consistent with the data obtained by some researchers [7, 20, 28, 31]. However, some authors have found these parameters to be dependent on age [16, 17]. What is interesting and surprising in group of men with normozoospermia, older men had slightly but significantly higher percentage of sperm progressive motility and vitality and those results highlighted how complicate could be estimation of age influence on basic semen parameters.
Age-dependent increase in sperm DNA fragmentation
To undertake more sophisticated evaluations of associations between male ageing and sperm quality, we performed DNA fragmentation as measured by the TUNEL (terminal deoxynucleotidyl transferase-mediated dUTP nick end labelling)/PI (propidium iodide) staining. In contrast to the comparison of standard semen parameters, in all groups, older men (≥40) had higher percentage of DFI that younger men. Several studies have presented similar data [18, 20, 23, 32]. The impact of age on sperm DNA integrity was revealed by Kaarouch et al. [20] and Alshahrani et al. [23], who showed that men ≥40 y had a significantly higher percentage of DFI than younger men. Similarly, Vagnini et al. [32] observed significant differences in the percentage of sperm with fragmented sperm DNA between patients ≤35 y vs. 36–39 y and vs. ≥40 y. Additionally, Plastira et al. [18] compared the sperm DNA integrity of men with oligoasthenoteratozoospermia and men with normozoospermia in two age-dependent groups: 24–34 y and 35–45 y. The researchers showed a significantly higher percentage of DFI in the group of older men with oligoasthenoteratozoospermia than the group of younger men. Moreover, they found significant correlations between age and the percentage of sperm cells with damaged chromatin. However, the authors did not find these differences or correlations in the group of men with normozoospermia. Similar results were obtained by Winkle et al. [19]; the group of men aged ≥40 y with abnormal standard semen parameters had a significantly higher percentage of DFI than subjects aged 36–39 y.
Based on Hallak [25], Borini et al. [26] and Benchaib et al. [27], we assumed that a >10% DFI is related to declining male fertility potential. The incidence of men with >10% DFI was higher in the group of subjects aged ≥40 y regardless of conventional characteristics of sperm. Moreover, older men had over double the risk of having >10% DFI compared to younger men.
Limitations of the study
Some limitations of our study should be addressed. Firstly, most of our subjects were in reproductive age, under 40 years and in all comparisons disproportion of groups size existed. In statistical analysis it is significant, and changes in groups proportions could influence the results. Secondly, we would like highlighted the fact that VitroLive Fertility Clinic and the Andrology Laboratory of Department of Histology and Developmental Biology are localized in the same city. Almost all of participants belong to the same nation and live in a similar environment. Because presented paper concern a global problem it would be valuable to performed multi-central research involving clinics and laboratories from other geographic regions. Thirdly, single assay was used to analyse the sperm DNA damage. DFI was verified only by TUNEL method, however this assay has strong clinical utility [25–27]. Further research with more DNA tests seems to be justified.
Final remarks
Our findings suggest a significant detrimental effect of advanced paternal age on sperm chromatin integrity because they reveal a significantly higher incidence of men with >10% DFI in the group of subjects aged ≥40 y. Furthermore, regardless of a standard semen analysis, groups of men after age 40 y are more than twice as likely to have >10% DFI than the groups of younger men and are recognized as having low fertility potential [25–27]. Therefore, our findings are consistent with the results of other authors reporting that a paternal age >40 y, commonly classified as advanced age, can be associated with a higher risk of reproductive failure [18-20, 23, 32]. It should be highlighted that the age-related changes are visible not only in selected conventional semen characteristics but also in the sperm chromatin status. It is particularly important because many authors have shown that the assessment of nuclear sperm DNA quality has greater clinical utility than standard semen analysis and better discriminates men with normal fertility potential from men with reduced fertility potential.
Methods
Study population
To perform this study, 1124 men (median: 33 y of age) were enrolled. The patients were partners in an infertile couples who were treated in the VitroLive Fertility Clinic (Szczecin, Poland) (n = 763) while volunteers with unknown fertility status (n = 361) reported to the Andrology Laboratory of Department of Histology and Developmental Biology (Pomeranian Medical University in Szczecin, Poland) for assessment of basic semen parameters. For all participants, the exclusion criteria included the following: azoospermia; a history of testicular torsion, cryptorchidism, testicular injury or cancer; varicocele; co-existing systemic disease; and a history of mumps. The ethics committee of the Pomeranian Medical University, Szczecin, Poland, approved this study protocol (ethical authorization number: KB-0012/10/14).
Standard semen analyses
The semen samples were obtained by masturbation after 2–7 days of sexual abstinence. The conventional semen parameters were assessed according to the World Health Organization 5th edition criteria [40] and performed in the Andrology Laboratory. Sperm motility (total, progressive and non-progressive motility) and vitality (eosin staining and hypoosmotic swelling test–HOS test) were determined under a contrast phase microscope and in bright field of light microscope, respectively (Primo Star, Zeiss, Germany). The sperm concentration was assessed with the improved Neubauer haemocytometer (Heinz Hernez Medizinalbedarf GmbH, Hamburg, Germany). The sperm morphology (including the TZI) was evaluated using the Papanicolaou staining method under a bright light microscope (CX 31 Olympus Optical Co., Ltd., Tokyo, Japan). Normozoospermia was considered according to the following criteria: sperm concentration ≥15 mln/mL, total number of sperm ≥39 mln, sperm progressive motility ≥32% and morphology ≥4%. Moreover, the TZI, vitality (eosin-negative and HOS-positive sperm cells) and concentration of round and peroxidase-positive cells (leukocytes) were evaluated. The standard semen evaluation was performed using a bright light microscope (CX31 Olympus Optical Co., Ltd., Tokyo, Japan). In the entire studied population the following seminological abnormalities of the standard sperm parameters were distinguished: normozoospermia (n = 448), asthenozoospermia (abnormal sperm motility, n = 107); asthenoteratozoospermia (abnormal sperm motility and morphology, n = 127); oligozoospermia (abnormal sperm number, n = 40); oligoasthenozoospermia (abnormal sperm number and motility, n = 31); oligoasthenoteratozoospermia (abnormal sperm number, motility and morphology, n = 133); oligoteratozoospermia (abnormal sperm number and morphology, n = 45) and teratozoospermia (abnormal sperm morphology, n = 193).
Evaluation of sperm DNA fragmentation index (DFI)
Nuclear DNA strand breaks in sperm cells were identified by TUNEL assay (terminal deoxynucleotidyl transferase-mediated dUTP nick end labelling)/PI (propidium iodide) assay using the FlowTACSTM Apoptosis Detection Kit (Trevigen, Inc., Gaithersburg, MD, USA) according to the manufacturer’s instructions. The liquefied ejaculate was centrifuged for 15 min at 300 g, and the pellet was washed twice in PBS (phosphate-buffered saline, Sigma Aldrich GmbH, St. Louis, USA) and fixed in 1% (v/v) formalin for 15 min at 4°C. After fixation, the pellet was washed twice (5 min, 300 g), resuspended with ice-cold 75% (v/v) ethanol and stored at -20°C for no less than two months. On the day of the test, samples were washed twice in PBS to remove the ethanol. Then, spermatozoa were permeabilized with Cytonin for 15 min at room temperature. After washing, the labelling reaction was performed.
The TUNEL test was based on the incorporation of substrate, biotin-labelled deoxynucleotide triphosphate (biotin-dNTP), into the free 3’-OH residues of the DNA fragments in the presence of terminal deoxynucleotidyl transferase (TdT). DNA breakage in the insertion sites of biotin-dNTP was identified by means of fluorescein isothiocyanate (FITC)-labelled streptavidin with a strong affinity for biotin. A negative control was obtained by omitting TdT from the reaction mixture, and a positive control was obtained by incubating sperm cells with DNase I. The TUNEL-positive spermatozoa (FITC-streptavidin-biotin-dNTP-labelled cells) were checked with a fluorescence microscope (BX41 Olympus Optical Co., Tokyo, Japan). After the TUNEL labelling reaction, previously permeabilized sperm cells (0.1% Triton X-100 in 0.1% sodium citrate) were stained with propidium iodide (PI) to discriminate apoptotic cells from necrotic cells in the flow cytometry analysis.
Flow cytometry measurements and data analysis
The verification of the incidence of sperm cells with SDF was performed using a Beckman Coulter flow cytometer (Cell LabQuanta SC MPL, Beckman Coulter, Fullerton, CA, USA) equipped with a 488 nm argon-ion laser. For each analysis, at least 10,000 events were collected at a flow rate of 150–250 events/s. The analysis of the results was performed using Cell LabQuanta SC MPL Analysis software (Beckman Coulter). The sperm population was gated on the basis of measurements of electronic volume (EV, parameter depends on cell size) and side scatter (SS, parameter depends on cellular granules). The green (480–550) and red (590–670) fluorescence signals were measured simultaneously using the FL1 and FL3 channels, respectively. The fluorescent data were obtained at a fixed gain setting in logarithmic (FL1, FL3) mode.
TUNEL/PI data analysis was performed according to the manufacturer’s instructions. The analyser threshold in the EV channel was adjusted to exclude debris and sperm aggregates from the flow cytometry analysis. The sperm-specific events were positively gated on the EV/SS dot plot. The calculation of TUNEL-positive spermatozoa with fragmented nuclear DNA (emitting green fluorescence) was based on positive (with DNase I) and negative (without TdT) histograms obtained for PI-stained spermatozoa (emitting red fluorescence). The staining of sperm cells with PI was performed after their permeabilization, and the positive and negative controls were generated for flow cytometry analysis to 1) exclude necrotic cells with fragmented DNA and 2) set a threshold to separate apoptotic sperm cells with DNA damage from cells with normal DNA integrity (Figure 2). The threshold was translated into histograms of test samples. The total events exhibiting green fluorescent intensities higher than the set threshold were considered apoptotic TUNEL-positive spermatozoa and expressed as a percentage, while the events emitting green fluorescence below the threshold were considered TUNEL-negative sperm cells displaying only background fluorescence (Figure 3).
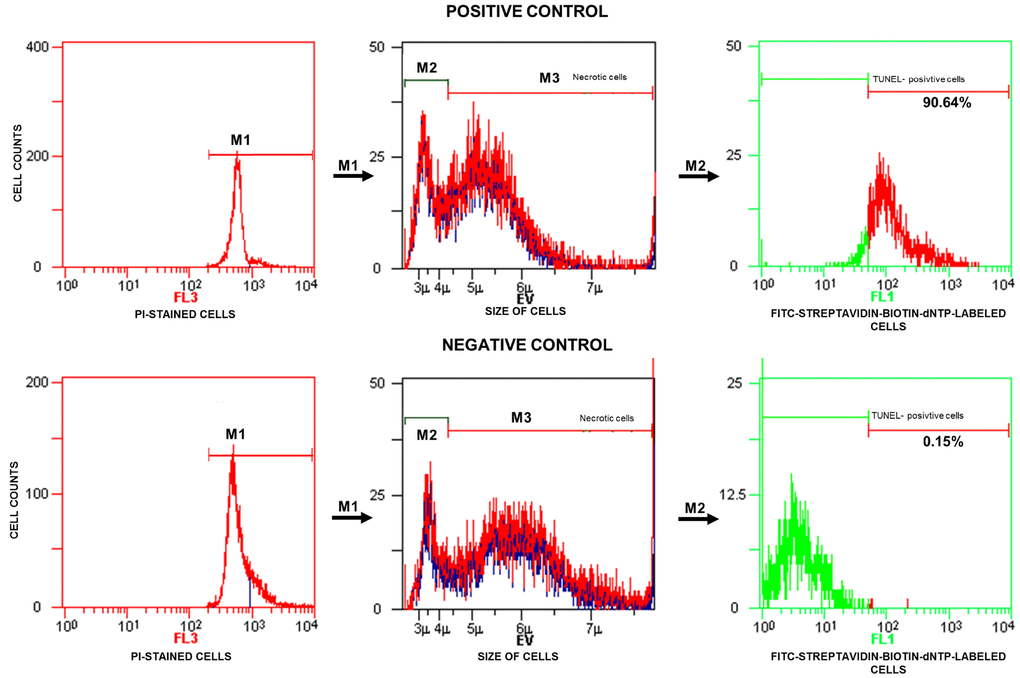
Figure 2. Representative histograms obtained for the positive (with DNase I) and negative (without TdT) controls and strategy applied in the TUNEL/PI flow cytometry analysis. In the fluorescence histograms, markers (M1, M2, M3) are set to exclude necrotic sperm cells and calculate the percentage of TUNEL-positive sperm cells (with fragmented DNA) read from the M2 marker. FL3 – red fluorescence channel for PI-stained cells, FL1 – green fluorescence channel for FITC-streptavidin-biotin-dNTP-labelled cells, TdT – terminal deoxynucleotidyl transferase, PI – propidium iodide, TdT – terminal deoxynucleotidyl transferase, TUNEL – terminal deoxynucleotidyl transferase-mediated dUTP nick end labelling.
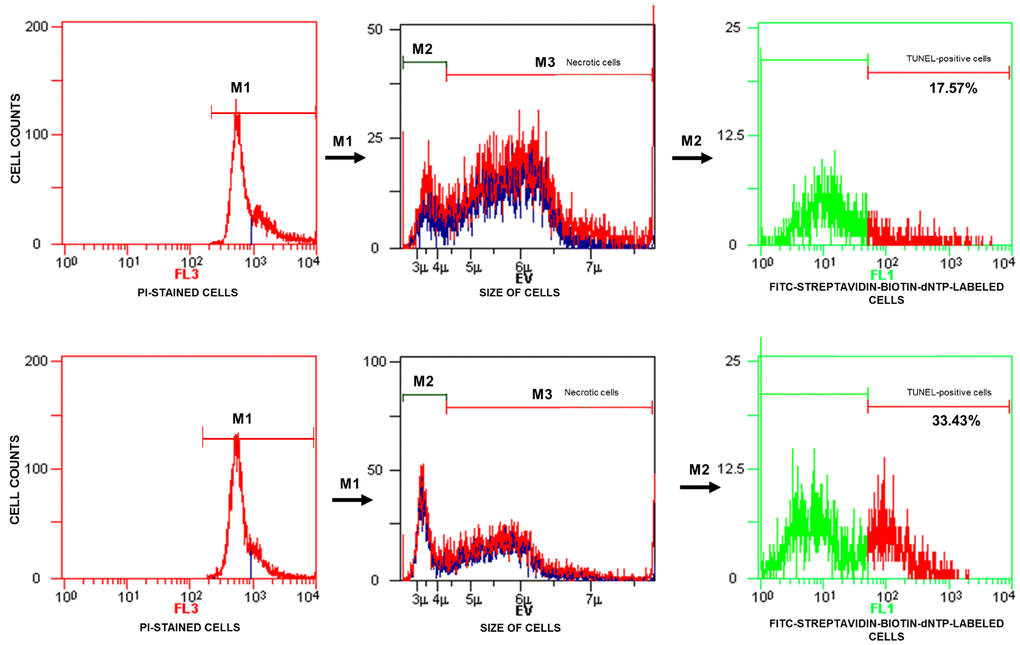
Figure 3. Representative histograms obtained for test samples. The calculation of TUNEL-positive sperm cells (with fragmented DNA) based on data analysis presented in Figure 1. FL3 – red fluorescence channel for PI-stained cells, FL3 – red fluorescence channel for PI-stained cells, FL1 – green fluorescence channel for FITC-streptavidin-biotin-dNTP-labelled cells, TUNEL – terminal deoxynucleotidyl transferase-mediated dUTP nick end labelling, PI – propidium iodide.
Statistical analysis
Statistical analysis was carried out using the software Statistica version 13.3 (StatSoft, Poland) and MedCalc version 15.2.2 (MedCalc Software, Belgium). Two independent group comparisons were performed using the Mann–Whitney U test for continuous variables. A chi-square test was used to compare the categorical data. The quantitative variables are expressed as the median (range) and means ± standard deviations (SDs), while qualitative data are presented as percentages. To define a risk for having high sperm DNA damage, an OR was calculated and is presented with the 95% confidence interval (CI) and p value. For all statistical tests, a p value of 0.05 was deemed significant.
Acknowledgments
We would like to thank VitroLive Fertility Clinic in Szczecin, Poland for the help in data acquisition.
Conflicts of Interest
The authors declare no conflicts of interest.
Funding
This study was supported by the Pomeranian Medical University in Szczecin, Szczecin, Poland (grants numbers: WNoZ-322-04/S/16/2019, MB-322-152/15).
References
- 1. Pan MM, Hockenberry MS, Kirby EW, Lipshultz LI. Male Infertility Diagnosis and Treatment in the Era of In Vitro Fertilization and Intracytoplasmic Sperm Injection. Med Clin North Am. 2018; 102:337–47. https://doi.org/10.1016/j.mcna.2017.10.008 [PubMed]
- 2. Uppangala S, Pudakalakatti S, D’souza F, Salian SR, Kalthur G, Kumar P, Atreya H, Adiga SK. Influence of sperm DNA damage on human preimplantation embryo metabolism. Reprod Biol. 2016; 16:234–41. https://doi.org/10.1016/j.repbio.2016.07.004 [PubMed]
- 3. Agarwal A, Mulgund A, Hamada A, Chyatte MR. A unique view on male infertility around the globe. Reprod Biol Endocrinol. 2015; 13:37. https://doi.org/10.1186/s12958-015-0032-1 [PubMed]
- 4. Levine H, Jørgensen N, Martino-Andrade A, Mendiola J, Weksler-Derri D, Mindlis I, Pinotti R, Swan SH. Temporal trends in sperm count: a systematic review and meta-regression analysis. Hum Reprod Update. 2017; 23:646–59. https://doi.org/10.1093/humupd/dmx022 [PubMed]
- 5. Khandwala YS, Zhang CA, Lu Y, Eisenberg ML. The age of fathers in the USA is rising: an analysis of 168 867 480 births from 1972 to 2015. Hum Reprod. 2017; 32:2110–16. https://doi.org/10.1093/humrep/dex267 [PubMed]
- 6. Rosiak A, Gill K, Jakubik J, Kups M, Patorski L, Kurzawa R, Piasecka M. [Czy zaawansowany wiek ojcowski ma wpływ na sukces rozrodczy? Część I: ocena wybranych parametrów seminologicznych] Is Advanced Paternal Age a Reproductive Risk? Part I: Assessment of Selected Standard Sperm Characteristics. Post Androl Online. 2017; 4:23–32.
- 7. García-Ferreyra J, Luna D, Villegas L, Romero R, Zavala P, Hilario R, Dueñas-Chacón J. High Aneuploidy Rates Observed in Embryos Derived from Donated Oocytes are Related to Male Aging and High Percentages of Sperm DNA Fragmentation. Clin Med insights Reprod Heal. 2015; 9:21–7. https://doi.org/10.4137/CMRH.S32769 [PubMed]
- 8. Belloc S, Hazout A, Zini A, Merviel P, Cabry R, Chahine H, Copin H, Benkhalifa M. How to overcome male infertility after 40: influence of paternal age on fertility. Maturitas. 2014; 78:22–29. https://doi.org/10.1016/j.maturitas.2014.02.011 [PubMed]
- 9. Rosiak A, Gill K, Jakubik J, Patorski Ł, Kurzawa R, Piasecka M. Is advanced paternal age a reproductive risk? Part II: embryo development, achieve pregnancy and health of offspring. Post Androl Online. 2017; 4:33–40.
- 10. Menezo Y, Servy E, Cohen M, Clement P. Advanced Paternal age and Endocrine Disruptors: Two causes of Psychiatric Disorders in Children, with DNA Methylation Dys-Regulation as a Common Biochemical Mechanism. In Psychiatric Disorders. 2017; 20:1–12.
- 11. Kimura R, Yoshizaki K, Osumi N. Risk of neurodevelopmental disease by paternal aging: A possible influence of epigenetic alteration in sperm. Adv Exp Med Biol. 2018; 1012:75–81. https://doi.org/10.1007/978-981-10-5526-3_8 [PubMed]
- 12. Conti SL, Eisenberg ML. Paternal aging and increased risk of congenital disease, psychiatric disorders, and cancer. Asian J Androl. 2016; 18:420–24. https://doi.org/10.4103/1008-682X.175097 [PubMed]
- 13. Sharma R, Agarwal A, Rohra VK, Assidi M, Abu-Elmagd M, Turki RF. Effects of increased paternal age on sperm quality, reproductive outcome and associated epigenetic risks to offspring. Reprod Biol Endocrinol. 2015; 13:35. https://doi.org/10.1186/s12958-015-0028-x [PubMed]
- 14. Lowe X, Eskenazi B, Nelson DO, Kidd S, Alme A, Wyrobek AJ. Frequency of XY sperm increases with age in fathers of boys with Klinefelter syndrome. Am J Hum Genet. 2001; 69:1046–54. https://doi.org/10.1086/323763 [PubMed]
- 15. Ramasamy R, Chiba K, Butler P, Lamb DJ. Male biological clock: a critical analysis of advanced paternal age. Fertil Steril. 2015; 103:1402–06. https://doi.org/10.1016/j.fertnstert.2015.03.011 [PubMed]
- 16. Stone BA, Alex A, Werlin LB, Marrs RP. Age thresholds for changes in semen parameters in men. Fertil Steril. 2013; 100:952–58. https://doi.org/10.1016/j.fertnstert.2013.05.046 [PubMed]
- 17. Cocuzza M, Athayde KS, Agarwal A, Sharma R, Pagani R, Lucon AM, Srougi M, Hallak J. Age-related increase of reactive oxygen species in neat semen in healthy fertile men. Urology. 2008; 71:490–94. https://doi.org/10.1016/j.urology.2007.11.041 [PubMed]
- 18. Plastira K, Msaouel P, Angelopoulou R, Zanioti K, Plastiras A, Pothos A, Bolaris S, Paparisteidis N, Mantas D. The effects of age on DNA fragmentation, chromatin packaging and conventional semen parameters in spermatozoa of oligoasthenoteratozoospermic patients. J Assist Reprod Genet. 2007; 24:437–43. https://doi.org/10.1007/s10815-007-9162-5 [PubMed]
- 19. Winkle T, Rosenbusch B, Gagsteiger F, Paiss T, Zoller N. The correlation between male age, sperm quality and sperm DNA fragmentation in 320 men attending a fertility center. J Assist Reprod Genet. 2009; 26:41–46. https://doi.org/10.1007/s10815-008-9277-3 [PubMed]
- 20. Kaarouch I, Bouamoud N, Madkour A, Louanjli N, Saadani B, Assou S, Aboulmaouahib S, Amzazi S, Copin H, Benkhalifa M, Sefrioui O. Paternal age: negative impact on sperm genome decays and IVF outcomes after 40 years. Mol Reprod Dev. 2018; 85:271–80. https://doi.org/10.1002/mrd.22963 [PubMed]
- 21. Urhoj SK, Andersen PK, Mortensen LH, Davey Smith G, Nybo Andersen AM. Advanced paternal age and stillbirth rate: a nationwide register-based cohort study of 944,031 pregnancies in Denmark. Eur J Epidemiol. 2017; 32:227–34. https://doi.org/10.1007/s10654-017-0237-z [PubMed]
- 22. Belloc S, Benkhalifa M, Cohen-Bacrie M, Dalleac A, Amar E, Zini A. Sperm deoxyribonucleic acid damage in normozoospermic men is related to age and sperm progressive motility. Fertil Steril. 2014; 101:1588–93. https://doi.org/10.1016/j.fertnstert.2014.02.006 [PubMed]
- 23. Alshahrani S, Agarwal A, Assidi M, Abuzenadah AM, Durairajanayagam D, Ayaz A, Sharma R, Sabanegh E. Infertile men older than 40 years are at higher risk of sperm DNA damage. Reprod Biol Endocrinol. 2014; 12:103. https://doi.org/10.1186/1477-7827-12-103 [PubMed]
- 24. Das M, Al-Hathal N, San-Gabriel M, Phillips S, Kadoch IJ, Bissonnette F, Holzer H, Zini A. High prevalence of isolated sperm DNA damage in infertile men with advanced paternal age. J Assist Reprod Genet. 2013; 30:843–48. https://doi.org/10.1007/s10815-013-0015-0 [PubMed]
- 25. Hallak J. Utility of sperm DNA fragmentation testing in different clinical scenarios of male reproductive abnormalities and its influence in natural and assisted reproduction. Transl Androl Urol. 2017 (Suppl 4); 6:S509–12. https://doi.org/10.21037/tau.2017.06.29 [PubMed]
- 26. Borini A, Tarozzi N, Bizzaro D, Bonu MA, Fava L, Flamigni C, Coticchio G. Sperm DNA fragmentation: paternal effect on early post-implantation embryo development in ART. Hum Reprod. 2006; 21:2876–81. https://doi.org/10.1093/humrep/del251 [PubMed]
- 27. Benchaib M, Braun V, Lornage J, Hadj S, Salle B, Lejeune H, Guérin JF. Sperm DNA fragmentation decreases the pregnancy rate in an assisted reproductive technique. Hum Reprod. 2003; 18:1023–28. https://doi.org/10.1093/humrep/deg228 [PubMed]
- 28. Whitcomb BW, Turzanski-Fortner R, Richter KS, Kipersztok S, Stillman RJ, Levy MJ, Levens ED. Contribution of male age to outcomes in assisted reproductive technologies. Fertil Steril. 2011; 95:147–51. https://doi.org/10.1016/j.fertnstert.2010.06.039 [PubMed]
- 29. Molina RI, Martini AC, Tissera A, Olmedo J, Senestrari D, de Cuneo MF, Ruiz RD. Semen quality and aging: analysis of 9.168 samples in Cordoba. Argentina. Arch Esp Urol. 2010; 63:214–22. https://doi.org/10.4321/s0004-06142010000300007 [PubMed]
- 30. Ng KK, Donat R, Chan L, Lalak A, Di Pierro I, Handelsman DJ. Sperm output of older men. Hum Reprod. 2004; 19:1811–15. https://doi.org/10.1093/humrep/deh315 [PubMed]
- 31. Park YS, Park S, Ko DS, Park DW, Seo JT, Yang KM. Observation of sperm-head vacuoles and sperm morphology under light microscope. Clin Exp Reprod Med. 2014; 41:132–36. https://doi.org/10.5653/cerm.2014.41.3.132 [PubMed]
- 32. Vagnini L, Baruffi RL, Mauri AL, Petersen CG, Massaro FC, Pontes A, Oliveira JB, Franco JG
Jr . The effects of male age on sperm DNA damage in an infertile population. Reprod Biomed Online. 2007; 15:514–19. https://doi.org/10.1016/S1472-6483(10)60382-3 [PubMed] - 33. Johnson SL, Dunleavy J, Gemmell NJ, Nakagawa S. Consistent age-dependent declines in human semen quality: a systematic review and meta-analysis. Ageing Res Rev. 2015; 19:22–33. https://doi.org/10.1016/j.arr.2014.10.007 [PubMed]
- 34. Zitzmann M. Effects of age on male fertility. Best Pract Res Clin Endocrinol Metab. 2013; 27:617–28. https://doi.org/10.1016/j.beem.2013.07.004 [PubMed]
- 35. Koh SA, Sanders K, Burton P. Effect of male age on oxidative stress markers in human semen. J Reprod Biotechnol Fertil. 2016; 5:1–10. https://doi.org/10.1177/2058915816673242
- 36. Asadi N, Bahmani M, Kheradmand A, Rafieian-Kopaei M. The impact of oxidative stress on testicular function and the role of antioxidants in improving it: A review. J Clin Diagn Res. 2017; 11:IE01–05. https://doi.org/10.7860/JCDR/2017/23927.9886 [PubMed]
- 37. Liguori I, Russo G, Curcio F, Bulli G, Aran L, Della-Morte D, Gargiulo G, Testa G, Cacciatore F, Bonaduce D, Abete P. Oxidative stress, aging, and diseases. Clin Interv Aging. 2018; 13:757–72. https://doi.org/10.2147/CIA.S158513 [PubMed]
- 38. Owyong M, Ramasamy R. Epidemiology of Oxidative Stress in Male Fertility. Oxidants, Antioxidants and Impact of the Oxidative Status in Male Reproduction. London: Academic Press; 2019. pp. 81–90. https://doi.org/10.1016/B978-0-12-812501-4.00009-2
- 39. Sabeti P, Pourmasumi S, Rahiminia T, Akyash F, Talebi AR. Etiologies of sperm oxidative stress. Int J Reprod Biomed (Yazd). 2016; 14:231–40. https://doi.org/10.29252/ijrm.14.4.231 [PubMed]
- 40. World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen. ed. Geneva: World Health Organization Press; 2010.