



Introduction
Biomarkers of aging are biological parameters that change in a predictable direction with aging in most individuals and, when assessed early in life, may predict subsequent longevity better than chronological age alone. Beyond their prognostic utility, the discovery of biomarkers of aging is attractive because they may shed light into the intrinsic mechanism of aging as a biological process [1]. Identifying biomarkers of aging may also provide insight into the biological mechanisms that accelerate or decelerate aging [2]]. Such biomarkers may be useful clinically for identifying persons at risk of developing adverse health outcomes traditionally associated with accelerated aging and to track the effectiveness of interventions aimed at slowing down the rate of aging and preventing its consequences such as multi-morbidity and disability.
miRNAs have emerged as important regulators of biological mechanisms that are relevant for aging. miRNAs are short non-coding RNAs that regulate gene expression generally by triggering mRNA decay and/or translational repression [3]. With over 1800 human miRNAs reported [4], miRNAs influence a wide range of biological functions, such as stem cell self-renewal, cell proliferation, apoptosis and metabolism [3].
Profiles of miRNAs found in plasma and serum have been linked to numerous cancers [5-8], cognitive impairment [9], Alzheimer’s disease [10,11] and other neurodegenerative disorders [12], and other pathologies [13], indicating that miRNAs are a new class of biomarkers of human diseases present in blood [14]. Because of the close relationship between these diseases and longevity, miRNAs may also serve as biomarkers of human aging. Our prior work has shown that miRNAs can serve as genetic biomarkers of aging in the nematode C. elegans [15]. Because miRNAs and aging genetic pathways are conserved from nematodes to humans, an increasing number of human miRNA studies have been carried out over the past several years. These studies have shown differential abundance of multiple miRNAs in peripheral blood mononuclear cells (PBMCs) or serum/plasma when comparing younger and older adults [16-21]. Sredni et al. found that changes in global miRNA levels, but not in mRNA levels, are associated with healthy aging in young adult women [16]. Noren Hooten et al. identified nine miRNAs that were differentially expressed in 30 vs. 64-year-old participants [17]. A summary of previous studies that have profiled miRNAs in biological samples of aged participants is provided in Table 1. These studies vary in the types of samples used, groups of participants, methods of profiling miRNA expression, and number of miRNAs profiled. Perhaps because of these differences, there is no overlap in the identified miRNAs that are up- or downregulated in the older vs. younger participants. More importantly, because these studies were based on case-control or cross-sectional designs, the lifespans of participants were not known and longitudinal blood samples were not analyzed.
Table 1. Related works on miRNAs and human aging samples, 2010-present
| Study | Samples | Analysis method | Participants' ages when miRNA expression was assessed | Main findings * | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Sredni et al. | Whole blood | Illumina 96-sample Universal Matrix Array (739 miRNAs) | 13 22-25 year olds (mean, 23.6 years) and 9 36-39 year olds (mean, 37.2 years) | The changes in global microRNA expression are associated with normal aging; the most differentially expressed microRNAs included miR-155, 8a, 142, 340, 363, 195, and 24 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Noren Hooten et al. | PBMCs | Multiplex qRT-PCR (over 800 miRNAs total) | 2 30 year olds and 2 64 year olds (male); validation in 14 young (mean, 30.1 years) and 14 old (mean, 64.2 years) individuals | 9 miRNAs downregulated (miR-24, 103, 107, 128, 130a, 155, 221, 496, 1538) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Gombar et al. | B cells | Deep sequencing (284 miRNAs); qRT-PCR (validation) | 3 63 year olds and 3 centenarians (female); validation in 27 individuals aged 50 to 100 years | 22 miRNAs upregulated, 2 downregulated; miR-363 downregulated (validation) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ElSharawy et al. | Whole blood | Microarray (863 miRNAs); qRT-PCR (7 miRNAs, validation) | 55 46 year olds and 15 centenarians and nonagenarians; validation in 17 younger (mean, 36.9 years) and 15 long-lived (mean, 101.5 years) individuals | 16 miRNAs upregulated, 64 downregulated; 3 miRNAs (miR-106a, 126, 30d) downregulated (validation) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Serna et al. | PBMCs | Microarray (1105 miRNAs) | 20 centenarians, 16 octogenarians, 14 young individuals | 6 miRNAs upregulated (centenarians vs. young) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Olivieri et al. | Plasma | ABI TaqMan miRNA PCR array (365 miRNAs); qRT-PCR (validation) | 11 20, 80, and 100 year olds; validation (only miR-21) in 111 healthy adults aged 20-105 (profiling cohort) and in 34 patients (mean, 87 years) with cardiovascular disease and 15 healthy centenarian offspring (mean, 72 years) | 46 miRNAs downregulated, 12 up- then downregulated, 5 upregulated in profiling cohort; one miRNA (miR-21) downregulated in validation cohort | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Noren Hooten et al. | Serum | Deep sequencing and qRT-PCR | 20 young (mean, 30.1 years) and 20 old (mean, 64.2 years) individuals | 3 miRNAs downregulated (miR-151a-5p, miR-181a-5p and miR-1248) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PBMCs, peripheral blood mononuclear cells. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| * The findings were made from the perspective of the long-lived participants, such as centenarians. There was no overlap in findings between all 7 studies. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
To address these limitations, we used miRNA PCR arrays to measure miRNA levels in serum samples obtained longitudinally at ages 50, 55, and 60 from 16 participants of the Baltimore Longitudinal Study of Aging (BLSA) who had documented lifespans. We compared miRNA expression changes not only across (i.e., between older and younger participants) but also within participants (using the three samples taken at different ages from each individual). In accordance with recent research that found a strong association between circulating miRNAs and human aging [22], our study suggests that circulating miRNAs are biomarkers of longevity.
Results
Basic characteristics of the study participants
As shown in Table 2, the 16 participants were all non-Hispanic males who were non-smokers. Their lifespans ranged from 58 to 92 years, and the years of death ranged from 1998 to 2008. The majority of participants died of heart disease or cancer. The range of lifespans followed a sigmoidal curve similar to that of the 406 BLSA participants from which the sample was derived (data not shown). The average lifespan was 75.5 years, compared with 75.6 years for U.S. males from the United Nations’ 2005-2010 life tables [23]; 8 participants had above average lifespan (long-lived subgroup: 76-92 years) and 8 had below average lifespan (short-lived subgroup: 58-75 years).
Table 2. Information from Baltimore Longitudinal Study of Aging (BLSA) regarding 16 participants in pilot study
| No. | Visit number* | Age when sample was taken | Ethnicity | Smoking status | DOB | DOD | Lifespan | Cause of death (disease) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S1 | 12 | 53.1 | White, not Hispanic Origin | former | 1/20/1926 | 9/29/2004 | 78.7 | heart disease | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S1 | 14 | 57.2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S1 | 17 | 63.1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S2 | 4 | 51.1 | White, not Hispanic Origin | former | 12/23/1912 | 12/11/2004 | 92.0 | cancer | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S2 | 6 | 54.6 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S2 | 9 | 61.4 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S3 | 8 | 54.6 | White, not Hispanic Origin | former | 10/31/1917 | 2/18/2004 | 86.3 | kidney neoplasm | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S3 | 10 | 58.3 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S3 | 13 | 64.0 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S4 | 11 | 50.4 | White, not Hispanic Origin | former | 1/13/1929 | 5/4/2004 | 75.3 | heart disease | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S4 | 13 | 54.3 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S4 | 16 | 60.3 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S5 | 8 | 50.9 | White, not Hispanic Origin | former | 5/8/1924 | 9/14/2007 | 83.4 | circulatory system disease | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S5 | 11 | 57.1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S5 | 13 | 61.7 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S6 | 10 | 50.0 | White, not Hispanic Origin | former | 1/31/1931 | 1/13/2007 | 76.0 | respiratory system disease | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S6 | 12 | 54.1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S6 | 15 | 60.3 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S7 | 7 | 51.0 | White, not Hispanic Origin | never | 2/27/1923 | 1/8/1998 | 74.9 | coronary heart disease | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S7 | 10 | 57.0 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S7 | 13 | 63.0 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S8 | 7 | 52.6 | White, not Hispanic Origin | former | 9/1/1920 | 1/22/2004 | 83.4 | nervous system disease | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S8 | 9 | 55.8 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S8 | 13 | 61.7 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S9 | 5 | 50.5 | White, not Hispanic Origin | former | 6/8/1922 | 12/22/2004 | 82.5 | nervous system disease | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S9 | 12 | 67.1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S9 | 13 | 69.3 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S10 | 13 | 50.1 | White, not Hispanic Origin | former | 10/13/1943 | 5/12/2001 | 57.6 | circulatory system disease | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S10 | 14 | 52.9 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S10 | 15 | 55.0 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S11 | 3 | 54.0 | White, not Hispanic Origin | former | 12/8/1918 | 1/3/2000 | 81.1 | neoplasm | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S11 | 9 | 70.0 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S11 | 10 | 72.5 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S12 | 9 | 53.2 | White, not Hispanic Origin | never | 5/8/1932 | 8/5/2003 | 71.2 | circulatory system disease | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S12 | 11 | 57.4 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S12 | 13 | 61.7 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S13 | 5 | 51.0 | White, not Hispanic Origin | former | 12/28/1924 | 2/11/1999 | 74.1 | cancer | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S13 | 7 | 55.0 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S13 | 10 | 61.0 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S14 | 10 | 52.0 | White, not Hispanic Origin | former | 8/14/1933 | 1/10/1998 | 64.4 | cancer | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S14 | 12 | 56.0 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S14 | 14 | 60.0 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S15 | 3 | 52.2 | White, not Hispanic Origin | former | 9/10/1935 | 12/6/1999 | 64.2 | circulatory system disease | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S15 | 4 | 57.7 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S15 | 7 | 63.1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S16 | 1 | 54.3 | White, not Hispanic Origin | former | 8/30/1934 | 10/31/2007 | 73.2 | neoplasm | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S16 | 3 | 58.9 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| S16 | 5 | 63.9 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DOB, date of birth; DOD, date of death. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| * Participants were followed for life with follow-up visits conducted at intervals of 1–4 years, depending on the participant's age, e.g., with visits approximately every 2 years for persons aged 60 or older. The 16 individuals (identified by number such as S1, S2, etc.) in the pilot study have at least three serum samples from around age 50, 55, and 60 available for analysis. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Magnitude of regulation and Pearson correlations
We compared the age 50 samples between the long-lived and short-lived subgroups by using methodology described in Figures S1 and S2. In total, we found 24 miRNAs that were significantly upregulated and 73 miRNAs that were significantly downregulated in long-lived participants. The 10 most upregulated and downregulated miRNAs are shown in Figure 1. The most upregulated miRNA was miR-373-5p, while the most downregulated miRNA was miR-15b-5p.
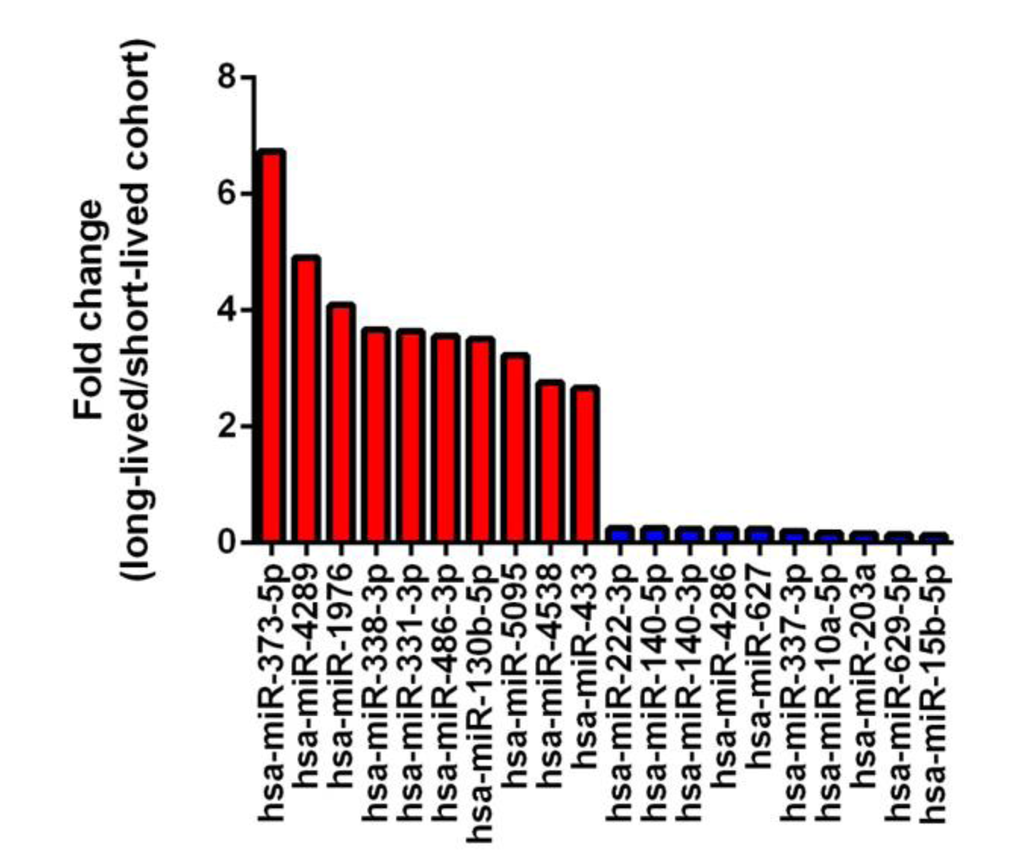
Figure 1. Fold change of 10 most up-regulated and down-regulated miRNAs (long-lived vs. short-lived subgroup)
We focused our analysis on comparing the longer-lived to the shorter-lived subgroup (Figure S1), as this gave more significant results than comparing the longest-lived quartile and shortest-lived quartile to the “average lifespan” subgroup (Figure S2) or analyzing intra-individual miRNA expression (Figure S3). Pearson correlations between miRNA expression in participants’ serum samples and participants’ lifespan were calculated, and the results of miRNAs that overlapped between the three different control methods (spike-in, global average, or stably-expressed miRNAs) are shown in Table 3. A total of nine miRNAs had correlations with p-values<0.05. miR-5095 and miR-378g also had more than one significant correlation per dataset. However, only six miRNAs (miR-211-5p, 374a-5p, 340-3p, 376c-3p, 5095, 1225-3p; in bold or italicized in Table 3) were correlated with lifespan and were significantly up or downregulated (above 2-fold or below 0.5-fold) when comparing subgroups of different lifespans, as shown in Figure S1 and S2. The correlations of these six miRNAs were reproducible in two separate experiments (data not shown). miR-340-3p had the highest correlation (all R2 values rounded up to two decimal places). miR-211-5p, miR-5095, and miR-1225-3p all had positive correlations and were similarly upregulated in the longer-lived subgroup but downregulated in the shorter lived subgroup. miR-374a-5p, miR-340-3p, and miR-376c-3p all had negative correlations and were similarly downregulated in the longer-lived subgroup but upregulated in the shorter lived subgroup. To illustrate these findings, Figure S4 shows an example of the correlation plot between the expression of miR-211-5p and lifespan for the age 50 subgroup; similar correlation plots were prepared for all six miRNAs.
Table 3. Pearson correlations between miRNA expression in participants’ serum samples and participants’ lifespan*
| miRNA | Spike-in Control | Global Average Control | Stable miRNAs Control | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Dataset † | R2 | p-value | Dataset † | R2 | p-value | Dataset † | R2 | p-value | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| hsa-miR-211-5p | 50 | 0.59 | 0.02 | 50 | 0.62 | 0.01 | 50 | 0.73 | 0.00 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| hsa-miR-29a-3p | 55 | 0.27 | 0.04 | 55 | 0.25 | 0.05 | (60-50)/50 | 0.44 | 0.01 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| hsa-miR-374a-5p | (60-50)/50 | 0.84 | 0.00 | (60-50)/50 | 0.78 | 0.01 | (60-50)/50 | 0.78 | 0.01 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| hsa-miR-340-3p ‡ | (55-50)/50 | 1.00 | 0.03 | (55-50)/50 | 1.00 | 0.03 | (55-50)/50 | 1.00 | 0.01 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| hsa-miR-376c-3p | 55 | 0.91 | 0.04 | 55 | 0.94 | 0.03 | 60 | 0.65 | 0.02 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| hsa-miR-5095 | 60 | 0.31 | 0.05 | 60-50 | 0.46 | 0.02 | 60 | 0.39 | 0.03 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| (60-50)/50 | 0.37 | 0.05 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| hsa-miR-1225-3p | 60 | 0.28 | 0.03 | 60 | 0.29 | 0.03 | 60 | 0.27 | 0.04 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| hsa-miR-3622a-5p | 55-50 | 0.37 | 0.03 | 55-50 | 0.41 | 0.03 | 60 | 0.29 | 0.04 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 60-50 | 0.37 | 0.03 | 60-50 | 0.46 | 0.01 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| hsa-miR-378g | 55-50 | 0.78 | 0.02 | 55-50 | 0.81 | 0.01 | 60 | 0.33 | 0.05 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 50 | 0.35 | 0.04 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| * Where p < 0.05 in all 3 control methods (p-values were rounded up to two decimal places). | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ‡ Italicized and underlined miRNA has perfect correlation, rounded up to two decimal places. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| † Dataset refers to the type of analysis of samples taken at different ages, such as around age 50 or age 60. The change in expression level between samples taken at different ages was also examined. For example, (55-50)/50 indicates that the Ct value of the sample around age 55 was subtracted by that of the sample around age 50, and the difference was divided by the Ct value of the sample around age 50. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Bioinformatics
After confirming that the six miRNAs of interest were found on the miRandola circulating miRNA database [24], we explored the aging pathways that these miRNAs might target. By using both miRTarBase and miRWalk, we identified validated targets of the candidate biomarker miRNAs; only miR-5095 did not have any validated targets. Table 4 includes 24 aging-associated mRNAs that are validated targets of miR-211-5p, 374a-5p, 340-3p, 376c-3p, and 1225-3p when checking for overlap with GenAge. We found that miR-1225 had the most aging-associated targets at 25%, in which 3 out of 12 validated targets were also listed in the GenAge database [25]. About 14% (8/56) of the miR-374a validated target mRNAs also encoded aging-associated proteins, as well as ~8% (1/12) of the miR-376c targets, ~7% (3/41) of the miR-211 targets, and ~7% (10/146) of the miR-340 targets. miR-340 had the most validated targets and the most aging-associated targets overall. One target, PARP1 mRNA [Poly(ADP-ribose) Polymerase 1], was found in the target lists of both miR-374a and miR-1225.
Table 4. Validated aging targets of five miRNAs from the pilot study
| miRNA * | Validated aging target mRNAs‡ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-211 | CREB5 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DDIT4 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IGF2R | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-340 | LMNA | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ARHGAP | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MPHOSPH | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IFG2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| YWHAZ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EEF1A1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| JUN | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PTEN | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CDKN2A | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HGF | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-374a | EP300 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ATM | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HMGB2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CISH | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PARP1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BCL2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TP73 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CDKN1A | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-376c | IGF1R | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-1225 | JUND | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PARP1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PRDX1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| * miR-5095 was not listed here because it did not have any validated aging targets. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ‡ miRTarBase + miRWalk, overlap with GenAge. Note that PARP1 (Poly(ADP-ribose) Polymerase 1) mRNA is a validated target of both miR-374a and miR-1225. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Discussion
We conducted a pilot study of miRNAs as biomarkers of aging by analyzing miRNA expression in serum samples from a longitudinal human aging study. We found that the expression of six circulating miRNAs in mid-adulthood significantly correlates with subsequent longevity, suggesting that these miRNAs may be useful biomarkers of human aging. As far as we know, this is the first study that directly correlates miRNA with human longevity using data from a longitudinal study.
Many interesting expression profiles were observed between study participants with different lifespans. For example, when comparing samples analyzed at age 50 between the long-lived and short-lived subgroups, we identified the 10 most differentially higher and lower expressed miRNAs (Figure 1). The most upregulated miRNA in long-lived participants, miR-373-5p, is part of the miR-373 family, which functions as a tumor suppressor in breast cancer [26]. The most downregulated miRNA in long-lived participants, miR-15b-5p, has been found to be upregulated in oral cancer cells [27]. Because lifespan is a complex trait characterized by escaping, delaying, or surviving fatal age-related diseases, including cancers, further scrutiny of the potential roles of the identified miRNAs in human aging is of great importance and interest.
The novel approach for the current study was to perform Pearson correlations between miRNA expression in all serum samples and the lifespan of 16 participants. Nine miRNAs had correlations with p values<0.05 (Table 3). Four of these miRNAs (miR-374, 376, 29, 378) have been shown in prior studies to be differentially expressed between older and younger persons in separate profiling experiments [18,20,21]. miR-374 was also downregulated in older participants in an earlier study [18], but in contrast with our study, miR-376 was upregulated in older participants in another report [20]. The trend in expression changes of the other two miRNAs (miR-29, 378) in prior studies was similar to our findings [20,21].
Six of the nine miRNAs (miR-211-5p, 374a-5p, 340-3p, 376c-3p, 5095, 1225-3p) may serve as useful biomarkers, as each of the six miRNAs were correlated with lifespan and were significantly up- or downregulated. Future studies can identify how examining expression of multiple miRNAs simultaneously versus one or a few miRNAs individually would affect these correlations. While some miRNA biomarker or disease-association studies have found significant correlations only by analyzing a profile of expression of multiple miRNAs [11,28], our study did identify miRNAs that individually correlate with lifespan. Further, it is striking that miRNA expression at ages 50, 55, and 60 correlates with the eventual, quite varied lifespans of the 16 participants in our pilot study.
Most of the 16 participants died from either heart disease or cancer (Table 2), and 15 participants were relatively healthy at the time of blood draw and only had coronary artery disease. The current study was not designed to evaluate the association between miRNA expression and disease-specific mortality. Nonetheless, our results suggest that these miRNAs may have separate functions during the process of aging itself. Interestingly, none of the six miRNAs has previously been shown to play a mechanistic role in aging, and none has been implicated in heart disease, but many have been shown to function and/or act as biomarkers in different cancers (miR-211: melanoma cell invasiveness; head, neck, renal cell carcinomas; pancreatic cancer; miR-374: small cell lung cancer; miR-340: osteosarcoma, colorectal cancer, breast cancer, gastric cancer; miR-376: glioblastoma, hepatocellular carcinoma) [24].
We identified 24 aging-associated mRNAs that are also validated targets of the five miRNAs (the sixth miRNA, miR-5095, did not have any validated targets). Notably, PARP1 mRNA, found in target lists of both miR-374a and miR-1225, encodes PARP1, a protein known for its role in repairing single-strand breaks during DNA replication [29,30]. PARP1 has also been linked to aging, being present in a complex with WRN DNA repair proteins that are deficient in participants with Werner syndrome, a premature aging syndrome [31][REMOVED HYPERLINK FIELD]. Additionally, one study found that there was higher PARP activity in cell lines established from blood samples of centenarians compared with younger participants [32]. PARP activity (due to PARP1) measured in blood samples of 13 mammalian species is also associated with maximum lifespan [33], suggesting an evolutionary role of PARP1 in determining species-specific lifespan. These findings indicate that PARP1 repair activity regulates mammalian longevity, consistent with the DNA damage theory of aging [34]. Additionally, both IGF1R mRNA (a miR-376c target) and IGF2R mRNA (a miR-211 target) encode receptors of the insulin signaling pathway (IGF1R and IGF2R, respectively), which has been tightly linked to variation in human longevity [35,36]. By examining aging-related roles of their genetic targets, we have identified a functional context for how these miRNAs might regulate aging processes. Additional research is needed to confirm correlations between the expression of these miRNAs and that of their target mRNAs, and to fully elucidate genetic regulation mechanisms within aging-related pathways.
Calculating Pearson correlations and separating participants into longer-lived vs. shorter-lived subgroups were only possible because we had obtained serum samples from a longitudinal study. This is a unique strength of the current study relative to prior miRNA profiling studies that have used blood samples from a single point in time among aged participants (Table 1). However, one limitation of our study is the relatively small sample size. Because this was a pilot study, we wanted to reduce the heterogeneity of our sample. This was accomplished through the use of several entry criteria (Figure 2), leading to the inclusion of 16 eligible men. Because of the small sample size, we did not account for multiple comparisons. Further validation of our results, especially in women and non-Whites, with larger sample sizes is needed.
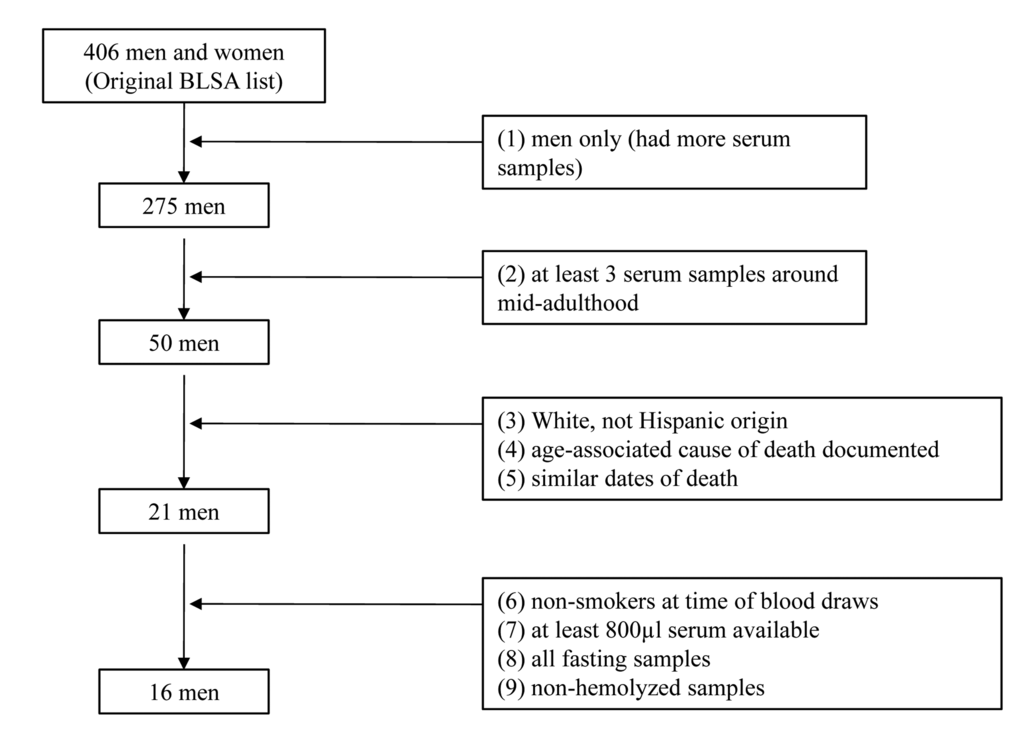
Figure 2. Flowchart showing the assembly of study participants for the pilot study
Our study has implemented a novel approach to identify human aging biomarkers in an attempt to translate basic scientific discoveries in model organisms to human aging. Our results suggest that the expression profiles of six miRNAs may be useful biomarkers of aging. Although preliminary, these results provide a basis for investigating miRNAs as potential predictors of future longevity, and they highlight the potential roles of select miRNAs in regulating aging processes, thus warranting further validation and mechanistic explorations.
Materials and Methods
Study design and setting
The Baltimore Longitudinal Study of Aging (BLSA) is a longitudinal study of human aging that began in 1958 with more than 1400 volunteers, ranging in age from 20s to 90s [37]. A detailed description of the BLSA has been provided previously [37,38]. Briefly, participants were assessed at the NIA Clinical Research Unit in Baltimore, Maryland by certified nurse practitioners and technicians following standardized protocols. The assessments included physiological parameters, biomarkers, risk factors, disease-related measures, impairments, and physical and cognitive function. Participants were followed for life with follow-up visits conducted at intervals of 1–4 years, depending on the participant's age, with visits approximately every 2 years for older persons aged 60 to 80 years and visits every year thereafter. All participants provided signed informed consent, and the BLSA protocol is approved by the NIEHS Institutional Review Board.
Participants
Longitudinal serum samples for this pilot study were obtained for 16 BLSA participants. The inclusion criteria were: male, availability of blood samples at around age 50 with at least three samples between ages 50 and 60, at least 800 µl of available serum, all fasting and non-hemolyzed samples (preventing introduction of miRNAs from red blood cells [39]), having a known cause of death, non-Hispanic White, and non-smoker (as miRNA dysregulation is linked to smoking-related diseases [40]) at the time of the blood draws. The assembly of participants for this study is provided in Figure 2. Eventually, we selected 16 participants (48 serum samples), all of whom had three blood samples taken within the range of ages 50, 55, and 60 (Figure S5). The sample was restricted to men because an inadequate number of women met the inclusion criteria.
miRNA extraction and profiling
The Serum/Plasma miRNA isolation kit, miScript II RT Kit (using HiSpec buffer), and miScript SYBR Green PCR Kit (Qiagen) were used to prepare samples for profiling following standard protocols. The miScript Human Serum & Plasma 384HC miRNA PCR Arrays (Qiagen) were used to profile 372 miRNAs and 12 controls (6 snoRNA/snRNA controls, a miRNA reverse transcription control, a positive PCR control, and a C. elegans miR-39 spike-in) for each of the 48 serum samples. A Roche LightCycler 480 machine was used to perform qPCR. Using the miScript miRNA PCR Array Handbook, the handbook protocol determined the same baseline and threshold across all amplification plots, in which the earliest amplification was manually set to cycle 15 and the threshold was set to 0.1. To confirm qRT-PCR results from the first RNA isolation, RNA was isolated a second time from serum in separate tubes from the same participants. In this second experiment, miScript Primer Assays were ordered to separately confirm expression of six identified miRNAs of interest (please see Ct analysis and Results section), along with spike-in control (miR-39) and four stable miRNAs (miR-21, 122, 126, 574) by following standard protocols.
Ct analysis
Data were analyzed using Sabio Sciences software to evaluate Ct values of controls before calculating fold changes. The miScript miRNA PCR Array Handbook advises that if the array is unbiased, normalize to the mean Ct of all expressed targets on the plate or to the mean Ct of at least four commonly expressed targets; however, we decided to perform both normalization methods, as well as normalizing to the spike-in control (see Data S1). The four miRNAs (miR-21, 122, 126, 574) with stable Ct values across all 48 serum samples were chosen from a list of stable miRNAs used for normalization by previous miRNA profiling studies using blood samples from aged participants [17-21,41,42]. The snoRNA and snRNA controls provided in each Qiagen miScript Human Serum & Plasma 384HC miRNA PCR Array were not used because the Ct values were all above 35 or were inconsistent on the same array. In the second experiment (referenced above), the mean Ct of all expressed targets was not used as a control due to using different assays (see Supplementary Material).
The method for calculating fold regulation for a given miRNA (repeated for each miRNA) among different age groups is described in detail in Figure S1 and S3. For example, comparing the long-lived with short-lived subgroup identified up- and downregulated miRNAs between the two subgroups (above 2-fold or below 0.5-fold). As shown in Figure S2, Pearson correlations were calculated using raw Ct values normalized to the appropriate controls (the number of controls varying depending on the experiment), and correlations were calculated between the Ct values and lifespans of the 16 participants. Only Pearson correlations that were statistically significant (p<0.05) using all controls were analyzed. Pearson correlations for each miRNA Ct value and the participant’s lifespan were calculated using Ct values from samples taken at different ages, including around age 50, 55, and 60. These calculations were repeated for 60-55 (the Ct value of the sample around age 60 was subtracted from that of the sample around age 55), 60-50, (55-50)/50 (the Ct value of the sample around age 55 was subtracted from that of the sample around age 50, and the difference was divided by the Ct value of the sample around age 50), and (60-50)/50. For each miRNA, any significant correlation from any of these Ct values was analyzed.
Bioinformatics
By using both miRTarBase and miRWalk, we identified validated targets of the candidate biomarker miRNAs. We then checked for overlap with a collection of 300 genes (300 mRNAs) listed in GenAge: The Ageing Gene Database from Human Ageing Genomic Resources [25,43] to determine if any of the mRNA targets of the remaining miRNAs might have aging-associated functions. The GenAge database includes a collection of genes associated with longevity and/or aging in model organisms (e.g., flies, mice) and humans.
Supplementary Materials
Conflicts of Interest
The authors have no conflict of interests to declare.
Funding
This research was supported by a grant to Dr. Slack from the NIH (AG033921) and by the Claude D. Pepper Older Americans Independence Center (P30AG021342). Dr. Gill is the recipient of an Academic Leadership Award (K07AG3587) from the National Institute on Aging.
References
- 1. López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. The hallmarks of aging. Cell. 2013; 153:1194–217. https://doi.org/10.1016/j.cell.2013.05.039 [PubMed]
- 2. Pincus Z, Slack FJ. Developmental biomarkers of aging in Caenorhabditis elegans. Dev Dyn. 2010; 239:1306–14. [PubMed]
- 3. Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004; 116:281–97. https://doi.org/10.1016/S0092-8674(04)00045-5 [PubMed]
- 4. Kozomara A, Griffiths-Jones S. miRBase: integrating microRNA annotation and deep-sequencing data. Nucleic Acids Res. 2011; 39:D152–57. https://doi.org/10.1093/nar/gkq1027 [PubMed]
- 5. Chen X, Ba Y, Ma L, Cai X, Yin Y, Wang K, Guo J, Zhang Y, Chen J, Guo X, Li Q, Li X, Wang W, et al. Characterization of microRNAs in serum: a novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008; 18:997–1006. https://doi.org/10.1038/cr.2008.282 [PubMed]
- 6. Mitchell PS, Parkin RK, Kroh EM, Fritz BR, Wyman SK, Pogosova-Agadjanyan EL, Peterson A, Noteboom J, O’Briant KC, Allen A, Lin DW, Urban N, Drescher CW, et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc Natl Acad Sci USA. 2008; 105:10513–18. https://doi.org/10.1073/pnas.0804549105 [PubMed]
- 7. Schetter AJ, Leung SY, Sohn JJ, Zanetti KA, Bowman ED, Yanaihara N, Yuen ST, Chan TL, Kwong DL, Au GK, Liu CG, Calin GA, Croce CM, Harris CC. MicroRNA expression profiles associated with prognosis and therapeutic outcome in colon adenocarcinoma. JAMA. 2008; 299:425–36. https://doi.org/10.1001/jama.299.4.425 [PubMed]
- 8. Yanaihara N, Caplen N, Bowman E, Seike M, Kumamoto K, Yi M, Stephens RM, Okamoto A, Yokota J, Tanaka T, Calin GA, Liu CG, Croce CM, Harris CC. Unique microRNA molecular profiles in lung cancer diagnosis and prognosis. Cancer Cell. 2006; 9:189–98. https://doi.org/10.1016/j.ccr.2006.01.025 [PubMed]
- 9. Sheinerman KS, Tsivinsky VG, Crawford F, Mullan MJ, Abdullah L, Umansky SR. Plasma microRNA biomarkers for detection of mild cognitive impairment. Aging (Albany NY). 2012; 4:590–605. https://doi.org/10.18632/aging.100486 [PubMed]
- 10. Geekiyanage H, Jicha GA, Nelson PT, Chan C. Blood serum miRNA: non-invasive biomarkers for Alzheimer’s disease. Exp Neurol. 2012; 235:491–96. https://doi.org/10.1016/j.expneurol.2011.11.026 [PubMed]
- 11. Leidinger P, Backes C, Deutscher S, Schmitt K, Mueller SC, Frese K, Haas J, Ruprecht K, Paul F, Stähler C, Lang CJ, Meder B, Bartfai T, et al. A blood based 12-miRNA signature of Alzheimer disease patients. Genome Biol. 2013; 14:R78. https://doi.org/10.1186/gb-2013-14-7-r78 [PubMed]
- 12. Gaughwin PM, Ciesla M, Lahiri N, Tabrizi SJ, Brundin P, Björkqvist M. Hsa-miR-34b is a plasma-stable microRNA that is elevated in pre-manifest Huntington’s disease. Hum Mol Genet. 2011; 20:2225–37. https://doi.org/10.1093/hmg/ddr111 [PubMed]
- 13. Gilad S, Meiri E, Yogev Y, Benjamin S, Lebanony D, Yerushalmi N, Benjamin H, Kushnir M, Cholakh H, Melamed N, Bentwich Z, Hod M, Goren Y, Chajut A. Serum microRNAs are promising novel biomarkers. PLoS One. 2008; 3:e3148. https://doi.org/10.1371/journal.pone.0003148 [PubMed]
- 14. Wang Y, Liang Y, Lu Q. MicroRNA epigenetic alterations: predicting biomarkers and therapeutic targets in human diseases. Clin Genet. 2008; 74:307–15. https://doi.org/10.1111/j.1399-0004.2008.01075.x [PubMed]
- 15. Pincus Z, Smith-Vikos T, Slack FJ. MicroRNA predictors of longevity in Caenorhabditis elegans. PLoS Genet. 2011; 7:e1002306. https://doi.org/10.1371/journal.pgen.1002306 [PubMed]
- 16. Sredni ST, Gadd S, Jafari N, Huang CC. A Parallel Study of mRNA and microRNA Profiling of Peripheral Blood in Young Adult Women. Front Genet. 2011; 2:49. https://doi.org/10.3389/fgene.2011.00049 [PubMed]
- 17. Noren Hooten N, Abdelmohsen K, Gorospe M, Ejiogu N, Zonderman AB, Evans MK. microRNA expression patterns reveal differential expression of target genes with age. PLoS One. 2010; 5:e10724. https://doi.org/10.1371/journal.pone.0010724 [PubMed]
- 18. ElSharawy A, Keller A, Flachsbart F, Wendschlag A, Jacobs G, Kefer N, Brefort T, Leidinger P, Backes C, Meese E, Schreiber S, Rosenstiel P, Franke A, Nebel A. Genome-wide miRNA signatures of human longevity. Aging Cell. 2012; 11:607–16. https://doi.org/10.1111/j.1474-9726.2012.00824.x [PubMed]
- 19. Gombar S, Jung HJ, Dong F, Calder B, Atzmon G, Barzilai N, Tian XL, Pothof J, Hoeijmakers JH, Campisi J, Vijg J, Suh Y. Comprehensive microRNA profiling in B-cells of human centenarians by massively parallel sequencing. BMC Genomics. 2012; 13:353. https://doi.org/10.1186/1471-2164-13-353 [PubMed]
- 20. Olivieri F, Spazzafumo L, Santini G, Lazzarini R, Albertini MC, Rippo MR, Galeazzi R, Abbatecola AM, Marcheselli F, Monti D, Ostan R, Cevenini E, Antonicelli R, et al. Age-related differences in the expression of circulating microRNAs: miR-21 as a new circulating marker of inflammaging. Mech Ageing Dev. 2012; 133:675–85. https://doi.org/10.1016/j.mad.2012.09.004 [PubMed]
- 21. Serna E, Gambini J, Borras C, Abdelaziz KM, Belenguer A, Sanchis P, Avellana JA, Rodriguez-Mañas L, Viña J. Centenarians, but not octogenarians, up-regulate the expression of microRNAs. Sci Rep. 2012; 2:961. https://doi.org/10.1038/srep00961 [PubMed]
- 22. Weilner S, Schraml E, Redl H, Grillari-Voglauer R, Grillari J. Secretion of microvesicular miRNAs in cellular and organismal aging. Exp Gerontol. 2013; 48:626–33. https://doi.org/10.1016/j.exger.2012.11.017 [PubMed]
- 23. Nations U. Department of Economic and Social Affairs, Population Division, World Population Prospects: The 2012 Revision. 2013.
- 24. Russo F, Di Bella S, Nigita G, Macca V, Laganà A, Giugno R, Pulvirenti A, Ferro A. miRandola: extracellular circulating microRNAs database. PLoS One. 2012; 7:e47786. https://doi.org/10.1371/journal.pone.0047786 [PubMed]
- 25. Hühne R, Thalheim T, Sühnel J. AgeFactDB--the JenAge Ageing Factor Database--towards data integration in ageing research. Nucleic Acids Res. 2014; 42:D892–96. https://doi.org/10.1093/nar/gkt1073 [PubMed]
- 26. Keklikoglou I, Koerner C, Schmidt C, Zhang JD, Heckmann D, Shavinskaya A, Allgayer H, Gückel B, Fehm T, Schneeweiss A, Sahin O, Wiemann S, Tschulena U. MicroRNA-520/373 family functions as a tumor suppressor in estrogen receptor negative breast cancer by targeting NF-κB and TGF-β signaling pathways. Oncogene. 2012; 31:4150–63. https://doi.org/10.1038/onc.2011.571 [PubMed]
- 27. Lu YC, Chen YJ, Wang HM, Tsai CY, Chen WH, Huang YC, Fan KH, Tsai CN, Huang SF, Kang CJ, Chang JT, Cheng AJ. Oncogenic function and early detection potential of miRNA-10b in oral cancer as identified by microRNA profiling. Cancer Prev Res (Phila). 2012; 5:665–74. https://doi.org/10.1158/1940-6207.CAPR-11-0358 [PubMed]
- 28. Xiao Y, Xu C, Guan J, Ping Y, Fan H, Li Y, Zhao H, Li X. Discovering dysfunction of multiple microRNAs cooperation in disease by a conserved microRNA co-expression network. PLoS One. 2012; 7:e32201. https://doi.org/10.1371/journal.pone.0032201 [PubMed]
- 29. Durkacz BW, Omidiji O, Gray DA, Shall S. (ADP-ribose)n participates in DNA excision repair. Nature. 1980; 283:593–96. https://doi.org/10.1038/283593a0 [PubMed]
- 30. Parsons JL, Dianova II, Allinson SL, Dianov GL. Poly(ADP-ribose) polymerase-1 protects excessive DNA strand breaks from deterioration during repair in human cell extracts. FEBS J. 2005; 272:2012–21. https://doi.org/10.1111/j.1742-4658.2005.04628.x [PubMed]
- 31. Mangerich A, Bürkle A. Pleiotropic cellular functions of PARP1 in longevity and aging: genome maintenance meets inflammation. Oxid Med Cell Longev. 2012; 2012:321653. https://doi.org/10.1155/2012/321653 [PubMed]
- 32. Muiras ML, Müller M, Schächter F, Bürkle A. Increased poly(ADP-ribose) polymerase activity in lymphoblastoid cell lines from centenarians. J Mol Med (Berl). 1998; 76:346–54. https://doi.org/10.1007/s001090050226 [PubMed]
- 33. Grube K, Bürkle A. Poly(ADP-ribose) polymerase activity in mononuclear leukocytes of 13 mammalian species correlates with species-specific life span. Proc Natl Acad Sci USA. 1992; 89:11759–63. https://doi.org/10.1073/pnas.89.24.11759 [PubMed]
- 34. Kenyon CJ. The genetics of ageing. Nature. 2010; 464:504–12. https://doi.org/10.1038/nature08980 [PubMed]
- 35. Pawlikowska L, Hu D, Huntsman S, Sung A, Chu C, Chen J, Joyner AH, Schork NJ, Hsueh WC, Reiner AP, Psaty BM, Atzmon G, Barzilai N, et al, and Study of Osteoporotic Fractures. Association of common genetic variation in the insulin/IGF1 signaling pathway with human longevity. Aging Cell. 2009; 8:460–72. https://doi.org/10.1111/j.1474-9726.2009.00493.x [PubMed]
- 36. Suh Y, Atzmon G, Cho MO, Hwang D, Liu B, Leahy DJ, Barzilai N, Cohen P. Functionally significant insulin-like growth factor I receptor mutations in centenarians. Proc Natl Acad Sci USA. 2008; 105:3438–42. https://doi.org/10.1073/pnas.0705467105 [PubMed]
- 37. Shock NW, Greulich RC, Andres R, Arenberg D. P.T. C, Jr., Lakatta EG and Tobin JD. (1984). Normal Human Aging: The Baltimore Longitudinal Study of Aging. (Washington, DC: US Government Printing Office: NIH Publication), pp. No. 84-2450.
- 38. Stone JL, Norris AH. Activities and attitudes of participants in the Baltimore longitudinal study. J Gerontol. 1966; 21:575–80. https://doi.org/10.1093/geronj/21.4.575 [PubMed]
- 39. Pritchard CC, Kroh E, Wood B, Arroyo JD, Dougherty KJ, Miyaji MM, Tait JF, Tewari M. Blood cell origin of circulating microRNAs: a cautionary note for cancer biomarker studies. Cancer Prev Res (Phila). 2012; 5:492–97. https://doi.org/10.1158/1940-6207.CAPR-11-0370 [PubMed]
- 40. Banerjee A, Luettich K. MicroRNAs as potential biomarkers of smoking-related diseases. Biomarkers Med. 2012; 6:671–84. https://doi.org/10.2217/bmm.12.50 [PubMed]
- 41. Olivieri F, Rippo MR, Procopio AD, Fazioli F. Circulating inflamma-miRs in aging and age-related diseases. Front Genet. 2013; 4:121. https://doi.org/10.3389/fgene.2013.00121 [PubMed]
- 42. Noren Hooten N, Fitzpatrick M, Wood WH
3rd , De S, Ejiogu N, Zhang Y, Mattison JA, Becker KG, Zonderman AB, Evans MK. Age-related changes in microRNA levels in serum. Aging (Albany NY). 2013; 5:725–40. https://doi.org/10.18632/aging.100603 [PubMed] - 43. Tacutu R, Budovsky A, Yanai H, Fraifeld VE. Molecular links between cellular senescence, longevity and age-related diseases - a systems biology perspective. Aging (Albany NY). 2011; 3:1178–91. https://doi.org/10.18632/aging.100413 [PubMed]