



Introduction
One of the most intriguing questions in stem cell biology is whether pluripotent stem cells (PSCs) exist in adult tissues. Several groups of investigators employing i) various isolation protocols, ii) detection of surface markers, and iii) experimental in vitro and in vivo models, have reported the presence of cells that possess a pluripotent character in adult tissues [1-4]. Such cells were assigned various operational abbreviations and names in the literature that added confusion to the field and raised the basic question of whether these are truly distinct or overlapping populations of the same primitive stem cells. Unfortunately, these cells were never characterized side-by-side to address this important issue. Nevertheless, taking into consideration their common features described in the literature, it is very likely that various investigators have described overlapping populations of developmentally early stem cells that are closely related [1-6].
From a developmental point of view, another important question is why should PSCs reside in adult organs? For many years it has been accepted that adult tissues contain only tissue-committed stem cells (TCSCs), such as epidermal stem cells, hematopoietic stem cells, or skeletal muscle stem cells, that have a limited potential for differentiation [7-11]. To address this question, we consider two scenarios that could occur during early embryogenesis and the development of lineage-restricted TCSCs [1,12,13]. In the first scenario, PSCs present in the inner cell mass of the blastocyst/epiblast, after giving rise to more differentiated lineage-restricted TCSCs, gradually disappear from the growing embryo and do not reside in adult tissues. In the second scenario, which we believe is more likely to take place during embryogenesis, some PSCs give rise to TCSCs but some survive in adult tissues as a backup population of PSCs that renews the pool of TCSCs over time. In this scenario, PSCs are precursors of TCSCs during organ/tissue rejuvenation and a source of these cells in emergency situations when organs are damaged (e.g., heart infarct or stroke). This scenario, however, requires such PSCs population deposited in adult tissues to be kept under control and in a quiescent state, which is essential to preventing uncontrolled proliferation leading to teratoma formation.
In this review, we will discuss the second scenario, in which population of very small embryonic- like stem cells (VSELs) - a rare population of epiblast-derived PSCs deposited in adult tissues is a reserve pool for TCSCs [1,14,15]. We will also present the most recent observations from our and other laboratories that support the presence of developmentally primitive stem cells in adult tissues with broad potential to differentiate into multiple lineages, which correspond to the VSELs described by our team.
Data supporting the presence of small stem cells in adult tissues
As mentioned above, several primitive cells with the characteristics of pluripotent or multipotent stem cells were isolated from adult murine and human tissues after expansion in in vitro cultures of cell suspensions isolated from murine and human bone marrow (BM) or human umbilical cord blood (UCB) or derived from adult organs after enzymatic processing by proteolytic enzymes [3-5,16,17]. In these in vitro cultures, in which cells grow while adhering to plastic or fibronectin, several populations of primitive cells were isolated, expanded and assigned different operational names [2,3,5,6,17-20]. Unfortunately, in none of these isolation procedures the phenotype of stem cell that initiated these cultures has been described clear from the beginning and the expanded in vitro cells were variously described as multipotent adult stem cells (MASCs) [19], unrestricted somatic stem cells (USSCs) [4,21,22] or marrow-isolated adult multilineage-inducible (MIAMI) cells [2].
In parallel, other isolation strategies were also employed, and an interesting population of small cells (ELH stem cells) able to differentiate into epithelial cells and hematopoietic cells was isolated from the murine BM by elutriation (E), lineage depletion (L), and the ability to home (H) to BM [23-25]. On the other hand, small cells able to differentiate into cells from all germ layers and called “spore-like stem cells” were isolated from adult mammalian tissues; however, the strategy by which they were purified and which surface markers they express was not provided in the original report [26].
Nevertheless, the presence of these primarily non-hematopoietic stem cells residing in adult BM challenged the concept of stem cell plasticity, which was supported by some data suggesting that hematopoietic stem cells (HSCs) may trans-differentiate into cells for non-hematopoietic tissues in murine models of experimental heart infarct, stroke, or liver damage [27,28]. The data in some of the experiments that showed a contribution of grafted BM cells into non-hematopoietic lineages gave the false impression that hematopoietic stem cells (HSCs) had trans-differentiated into cells for non-hematopoietic tissues [29,30].
Rejecting the concept that trans-differentiation of adult HSCs into non-hematopoietic lineages is a frequent biological phenomenon, our main aim became to isolate a population of true PSCs from BM, which could explain the data showing a contribution from BM cells in the regeneration of damaged organs. Based on preliminary data gained from our initial experiments, we purified from adult murine BM a population of cells slightly smaller than red blood cells that expressed the surface phenotype Sca1+Lin−CD45− and markers of pluripotent stem cells (e.g., Oct4, Nanog, and SSEA-4). These cells displayed a distinct morphology characterized by a high nuclear/cytoplasmic ratio and undifferentiated chromatin [31, 32]. Phenotypically similar cells were subsequently purified from murine fetal liver, as well as from several adult murine organs, including brain, skeletal muscle, kidney, lung, liver, and pancreas [33-36].
Small cells comprising population of human VSELs were also purified from neonatal umbilical cord blood (UCB) and adult patient-mobilized peripheral blood (mPB) [37-43]. Human UCB and mPB are enriched in VSELs within a population of CD133+Lin−CD45− cells that co-express CXCR4, and some of them are also CD34+. Like their murine counterparts, human UCB and mPB VSELs also express Oct4 and Nanog transcription factors in nuclei and express SSEA-1 on the surface [37-42]. The procedure for how to purify these cells has been described in detail in several publications and summarized in Current Protocols in Cytometry [44] and Methods in Cell Biology [14]. In this review, we will focus on new observations generated by our team, as well as the work of other groups working on VSELs or closely related cells.
Recent observations from our group on murine and human cells that are attributable to VSELs
In the last years we have put considerable effort into better characterizing these rare cells at the molecular level.
- Progress on VSEL isolation strategies
VSELs from human UCB and mPB were initially purified from an erythrocyte-depleted population of nucleated cells by multiparameter sorting as a population of small CD133+ CD45− Lin− cells [37]. However, this procedure is time consuming and the sorting time required to process an entire cord blood unit (~50-100 ml) to isolate rare VSELs from UCB mononucleated cells takes up to 3-4 days [37]. Therefore, from the beginning it has been clear that a more rapid and less-expensive method for isolation of these cells must be found. In order to establish a more efficient method for VSEL purification from UCB, we employed a three-step isolation strategy based on removal of erythrocytes by hypotonic lysis (1st step), followed by immunomagnetic separation of CD133+ cells (2nd step), followed by FACS-based isolation of CD133+Lin−CD45− cells (3rd step). The entire isolation procedure takes only 2-4 hours per 100 ml of UCB [39,44].
As an alternative strategy, we recently proposed to reduce the number of lineage factors required for purification by exposure of erythrocyte-depleted, immunomagnetic bead-selected CD133+ cells to Aldefluor, followed by staining with anti-CD133+ antibodies and only two lineage-specific antibodies (anti-CD45 and anti-GlyA) for step 3 [38]. Aldefluor is a substrate for aldehyde-dehydrogenase (ALDH), a cytosolic enzyme highly expressed in less-differentiated hematopoietic cells, and is implicated in resistance to some alkylating agents [60,61]. In the presence of ALDH, Aldefluor becomes modified to a fluorescent molecule that may then be used to mark ALDH-expressing cells. Inclusion of anti-GlyA antibody is based on the well-known fact that small erythroblast GlyA+ cells present in UCB do not express the CD45 antigen.Thus, selection for CD45− cells could enrich for these unwanted cells. By employing this strategy, we are able to subsequently sort CD133+ cells enriched for VSELs and are able to obtain ~103 CD133+/ CD45−/GlyA−/ALDHlowand 4x103 CD133+/ CD45−/GlyA−/ALDHhigh UCB-VSELs from 100 ml of UCB [38]. These numbers demonstrate how rare these cells are in UCB. When we compared both fractions of VSELs by RT-PCR gene expression analysis, we found that CD133+/CD45−/GlyA−/ALDHlowVSELshave a higher expression of Oct4 than the CD133+/CD45−/GlyA−/ALDHhigh fraction [38].
However, we are aware that there is yet room to improve sorting and to develop alternative strategies for purifying UCB-VSELs. To accomplish this goal, one should consider the use of metabolic fluorochromes to see whether VSELs are enriched among Rh123dull, Pyronin Ylow, and Hoechst 33342low cells. Moreover, we expect that our proteomics data analysis with UCB-VSELs will reveal the presence of new positive markers that could be employed for purification of these rare cells.
- Mobilization of VSELs into PB in response to organ/tissue damage
VSELs circulate in PB under steady-state conditions; however, the number of these cells is very low. In our recent work we provided evidence that VSELs can be mobilized into peripheral blood in adult patients injected with granulocyte-colony stimulating factor (G-CSF). This observation laid the foundation for the concept that G-CSF mobilization can be employed to harvest these cells from patients for therapeutic purposes.
Furthermore, our studies on VSEL mobilization into PB revealed that VSELs are mobilized not only, as described previously, in patients suffering from heart infarct [40,42,62] or stroke [41] but also in patients suffering from skin burns [63], active inflammatory bowel disease [64], and cancer [14]. More importantly, based on our preliminary data, we also envision that the number of VSELs circulating in PB in patients could be of prognostic value. This however, requires further study and long-term clinical correlations.
- Novel characteristics of human VSELs
In the past two years we investigated several experimental approaches to better characterizing murine and human UCB-derived VSELs, both at the molecular and morphological levels. First, our molecular gene array analysis studies on limited numbers of highly purified VSELs revealed that murine VSELs are somewhat heterogenous developmentally [65-68]. In particular, while all of them express the pluripotency marker Oct-4, some of them express genes that are more closely related to genes expressed by epiblast-derived stem cells and others to genes expressed by migrating primordial germ cells. This shows that VSELs resemble early stem cells developmentally; if isolated from adult BM, they may differ somewhat in their differentiation potential [65-68].
Based on well-established FACS identification protocols, we also found that CD133+Lin−CD45−VSELs identified in UCB (Figure 1 panel A), like their murine counterparts, i) highly express telomerase (not published), ii) are diploid (Figure 1 panel B), and iii) are viable, as shown by their ability to exclude the viability dye (7-aminoactinomycin D [7-AAD], Figure 1 panel C). Importantly, we observed that the highest number of pluripotent Oct-4+ VSELs reside in the population of CD133+Lin−CD45−UCB-derived cells [37,39]. Moreover, some of the CD133+Lin−CD45− VSELs, which represent only a very small subfraction among UCB Lin−CD45−non-hematopoietic cells, may co-express other stem cell markers, including CD34, CXCR4, and SSEA-4 [37,39]. Thus, VSELs represent a very rare subpopulation of UCB stem cells among Lin−CD45− non-hematopoietic UCB cells, which contains other stem cell types, including endothelial progenitor cells (EPCs) and mesenchymal stem cells (MSCs), and may be identified based on very small size (FSClow/SSClow) and co-expression of CD133, CD34, and, CXCR4 [37,39].
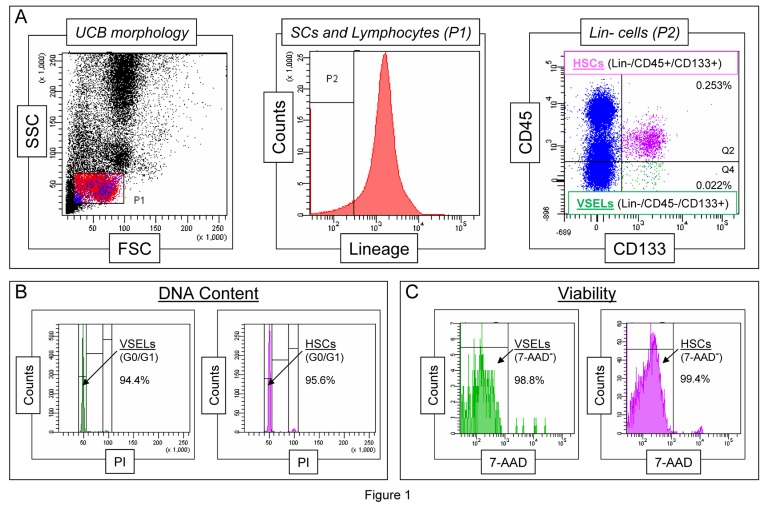
Figure 1. Morphological characteristics of UCB-derived VSELs by flow cytometric assays Panel A shows a well-established flow cytometric protocol for identification of CD133+Lin−CD45− VSELs among the whole-nucleated fraction of UCB cells. The dot-plot (left) visualizes cells based on FSC and SSC parameters, indicating the relative cellular size and complexity, respectively. The FSClow/SSClow lymphocytic population, which includes very small objects, is enclosed in region P1, and the cells were further analyzed according to hematopoietic lineage marker expression (middle histogram). Lin- cells from region P2 are plotted in the dot-plot representing CD133 and CD45 expression (right). VSELs are identified as CD133+Lin−CD45− cells (region Q4), while HSCs are identified as CD133+Lin-CD45+ cells (region Q2). Percentages indicate the content of both stem cell populations among all nucleated cells in one representative UCB sample. Panel B presents a representative analysis of DNA content in UCB-derived VSELs and HSCs following fixation and staining with propidium iodide (PI). The percentages indicate normal diploid (2n) VSEL and HSC fractions in the G0/G1 phase of the cell cycle. Panel C shows the viability of VSELs and HSCs examined by flow cytometry following the staining of freshly isolated cells with a viability dye, 7-aminoactinomycin D (7-AAD). Viable VSELs and HSCs are represented as 7-AAD− cells, which exclude the dye, and their percentage content is computed for each of the stem cell populations.
- In vitro and in vivo evidence that VSELs could be pluripotent stem cells
There are stringent criteria for classifying a stem cell as a PSC. In Table I are listed the most important in vitro and in vivo criteria that we expect from PSCs. These criteria were established by embryologists who are working with embryonic stem cells (ESCs) isolated from embryos or on established in vitro-immortalized ESC lines or induced pluripotent stem cells (iPSCs). However, some of these stringent criteria listed as in vivo criteria of pluripotency, such as complementation of blastocyst development and teratoma formation (Table I), are not always visible in pluripotent epiblast-derived stem cells [69,70].
Table I. In vitro and in vivo features of pluripotent stem cells (PSCs)
| In vitro criteria for PSCs | VSELs |
|---|---|
| 1. Undifferentiated morphology, euchromatin, and high nuclear/cytoplasm ratio 2. PSC markers (e.g., Oct-4, Nanog, and SSEA), open chromatin at the Oct-4 promoter, bivalent domains, and female PSCs reactivate the X chromosome 3. Multilineage differentiation into cells from all three germ layers (meso-, ecto-, and endoderm) | Yes Yes Yes |
| In vivo criteria for PSCs | VSELs |
| 1. Complementation of blastocyst development 2. Teratoma formation | No No |
Our most recent experimental data support the conclusion that murine VSELs fulfill all the in vitro criteria listed in Table I. In particular, murine VSELs not only possess the primitive morphology of early developmental cells but also express typical markers for PSCs (e.g., Oct-4, Nanog, and Rex-1) [31,33,34,37,71]. Significantly, we also recently confirmed the true expression of Oct-4 in murine VSELs (which was previously supported by the presence of an open-type chromatin in the Oct-4 promoter) by direct analysis of its methylation state [72] and association with transcription-permissive histones [65,66]. Specifically, we observed that the Oct-4 promoter in murine VSELs is hypomethylated, and by employing the carrier-ChIP assay, we found that the chromatin in the Oct-4 promoter is associated with the gene transcription-promoting histone H3Ac, while its association with the transcription-restricting histone H3K9me2 is relatively low [65,66,72]. We also evaluated the epigenetic state of another core transcription factor, Nanog, and observed that its promoter has a higher level of methylation in VSELs (~50%). In quantitative ChIP experiments performed in parallel, we also observed that the H3Ac/H3K9me2 ratio favors transcription and supports an active state [65,66,72]. Based on these results, we conclude that murine VSELs truly express both of the embryonic transcription factors Oct-4 and Nanog.
With respect to the other in vitro criteria of pluripotency (Table I), murine VSELs also possess bivalent domains in promoters that encode developmentally important homeobox-containing transcription factors (Sox21, Nkx2.2, Dlx1, Lbx1h, Hlxb9, Pax5, and HoxA3) [68]. Furthermore, VSELs derived from female mice reactivate the X-chromosome.
Finally, we succeeded in differentiation of VSELs in vitro into cells from all three germ layers, while recently published data demonstrated that murine BM-derived VSELs can be specified in vivo into HSCs [73], mesenchymal stem cells (MSCs) [49], and cardiomyocytes [74,75].
Our in vitro data on differentiation of human VSELs are limited at this point but so far we have shown that human UCB-derived VSELs, under appropriate culture conditions, can also be specified into the hematopoietic lineage [38].
However, taking into consideration the in vivo criteria expected from PSCs (Table I), murine VSELs do not complete blastocyst development and do not grow tera- tomas. This discrepancy between in vitro and in vivo criteria of PSCs for VSELs has recently been explained by epigenetic changes in expression of some paternally imprinted genes [67,72]. This phenomenon will be discussed below in the context of the developmental origin of VSELs and the mechanisms that keep them quiescent in adult tissues and prevent them from uncontrolled proliferation and teratoma formation [67,72].
Novel molecular evidence supporting the developmental origin of VSELs
As mentioned above, gene expression studies have revealed that VSELs express several epiblast and germ line markers and we hypothesize that VSELs originate from early epiblast-derived migrating primordial germ cell (PGC)-like cells, are deposited during development in adult tissues as a source of TCSCs, and play a role in organ rejuvenation [76,77]. In support of this notion, molecular analysis of murine BM-derived VSELs has revealed that these cells express several genes that are characteristic of epiblast SCs (Gbx2, Fgf5, and Nodal) and germ line specification (Stella, Prdm14, Fragilis, Blimp1, Nanos3, and Dnd1) [65,66,72].
However, a main goal of the molecular analysis studies was to explain why VSELs do not fulfill the in vivo gold-standard criteria expected for PSCs (complementation of blastocyst development and teratoma formation in immunodeficient animals), which are seen with ESCs and iPSCs. To explain this discrepancy, we observed that VSELs, in a similar manner as late migratory primordial germ cells (PGCs), modify the methylation of imprinted genes, preventing them from uncontrolled proliferation and explaining their quiescent state in adult tissues [65,66].
It is well known that imprinted genes play a crucial role in embryogenesis, fetal growth, the totipotential state of the zygote, and the pluripotency of developmentally early stem cells [78,79]. It has been demonstrated that VSELs freshly isolated from murine BM erase the paternally methylated imprints at regulatory differentially methylated regions (DMRs) within the Igf2-H19 and Rasgrf1 loci; however, they also hypermethylate the maternally methylated imprints at DMRs for the Igf2 receptor (Igf2R), Kcnq1-p57KIP2, and Peg1 loci [72].Because paternally expressed imprinted genes (Igf2 and Rasgrf1) enhance embryonic growth and maternally expressed genes (H19, p57KIP2, andIgf2R) inhibit cell proliferation [72], the unique genomic imprinting pattern observed in VSELs demonstrates the growth-repressive influence of imprinted genes on these cells [72].
All these results suggest that epigenetic reprogramming of genomic imprinting maintains the quiescence of Oct4+ epiblast/germ line-derived VSELs deposited in the adult body and protects them from premature ageing and uncontrolled proliferation (e.g., teratoma formation). On the other hand, reversal of this mechanism will be crucial to employing VSELs as a population of PSCs in regenerative medicine. Currently, we are testing whether downregulation of the expression of H19 enhances VSEL expansion, as has recently been demonstrated for PSCs derived by parthenogenesis [80,81].
The role of murine VSELs in tissue rejuvenation and longevity
Interestingly, our most recent work has revealed that murine VSELs modulate by epigenetic changes the expression of imprinted genes, such as Igf2-H19, RasGRF1, and IGF2R, that play an important a role in insulin/insulin-like growth factors signaling (IIS), as well as a crucial role in maintaining the pool of VSELs residing in adult tissues [82,83]. Accordingly, we observed that murine BM-sorted VSELs erase the paternally methylated imprints within the DMRs for Igf2-H19 and RasGrf1, while they hypermethylate the maternally methylated DMR forIgf2R. The epigenetic modification of imprinted loci (including Igf2-H19, RasGRF1, and IgfR) prevents VSELs from uncontrolled proliferation and teratoma formation. This epigenetic modification of imprinted genes explains why murine VSELs, despite expressing several markers of pluripotency (e.g., an open chromatin structure at the promoters for Oct-4 and Nanog), the presence of bivalent domains at developmentally important homeobox-domain containing genes, the reactivation of the X chromosome in female VSELs, and in vitro differentiation into cells from all three germ layers, do not complement blastocyst development after injection into the pre-implantation blastocyst (Table I).
To summarize, the abovementioned changes in expression of imprinted genes in murine VSELs lead to perturbation of IIS by downregulation of i) IGF2, which is an autocrine factor involved in proliferation of VSELs, and ii) RasGRF1, which is a GTP-exchange factor (GEF) crucial for signaling from activated IGF-IR and the insulin receptor (InsR, Figure 2). In addition, hyperemethylation of DMRs on the maternal chromosome encoding IGF-2R [83] has an additional negative affect on IIS in VSELs. As mentioned above, IGF2R serves as a decoy receptor that prevents IGF-2 from binding to IGF-IR [83]. This epigenetic reprogramming of genomic imprinting negatively affects IIS signaling, maintains the quiescent state of murine VSELs, and thus protects them from premature depletion from the tissues and prevents their involvement in tumor formation.
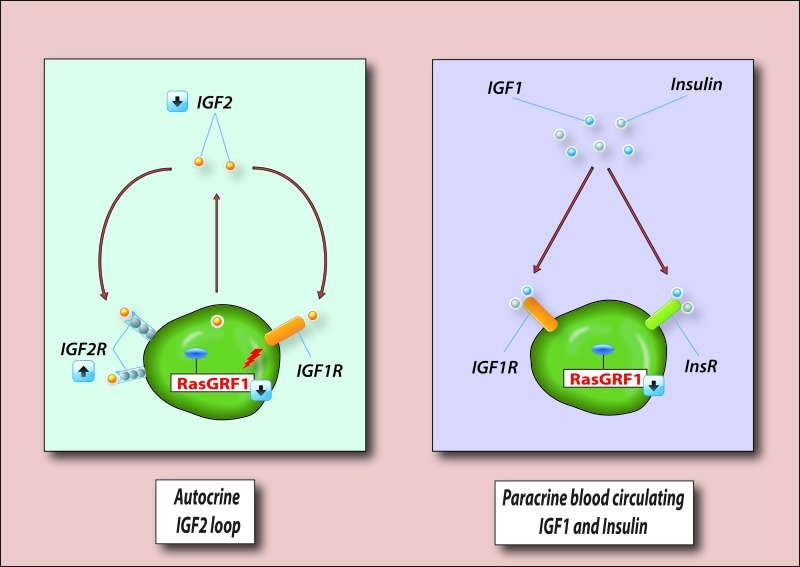
Figure 2. VSELs deposited in adult tissues are protected from excessive insulin/insulin-like growth factor signaling (IIS) VSELs are deposited in adult tissues as a backup population for tissue-committed stem cells (TCSCs) and are protected from IIS, which would otherwise lead to their premature depletion from adult tissues, as well as trigger uncontrolled proliferation and teratoma formation. Left panel - VSELs are protected from autocrine IIS by changes in expression of imprinted genes that are important in IIS. Downregulation of autocrine expression of IGF2, upregulation of IGF2R (a non-signaling receptor that binds IGF2 and prevents its binding to signaling IGF1R), and downregulation of RasGRF1 (which is involved in signal transduction from IGF2R), makes VSELs less sensitive to autocrine/paracrine IGF2 signaling. Right panel - Downregulation of RasGRF1, which is involved in signaling from activated IGF1R and InsR in VSELs, also plays an important role in attenuation of IIS signaling by paracrine circulating levels of IGF1 and insulin. However, in the presence of chronic elevated levels of IGF1 and insulin in blood, VSELs deposited in adult tissues may proliferate in an uncontrolled manner and become depleted much faster over time. This may contribute to the accelerated aging observed in situations with high circulating IGF1 and insulin levels (e.g., Laron dwarf mice, Ames dwarf mice, or chronic high caloric uptake). At the same time, chronic exposure to IIS may also trigger uncontrolled activation of VSELs and their malignant transformation. For reasons of simplicity, other imprinted genes not involved in IIS that negatively affect VSEL proliferation (H19 and p57Kip2) are not shown.
Based on these findings and published reports that elevated IIS signaling negatively affects life span in experimental animals [84], we proposed a hypothesis that relates aging, longevity, and IIS to the abundance and function of pluripotent VSELs deposited in adult murine tissues [82,83]. We postulate that a decrease in the number of these cells due to prolonged IIS negatively affects the pools of TCSCs in various organs and has an impact on tissue rejuvenation and life span [82,83]. In support of this notion, we observed a significantly higher number of VSELs in long-living murine strains (e.g., Laron dwarfs and Ames dwarfs), whose longevity is explained by low levels of circulating IGF1 and a decrease in IIS [85]. By contrast, the number of VSELs is reduced in mice with high levels of circulating IGF1 and enhanced IIS (e.g., growth hormone-overexpressing transgenic mice) compared to normally aging littermates [85].
The influence of IIS on the pool of VSELs residing in adult tissues seems to be twofold (Figure 2). First, since IGF2 is an autocrine factor for these cells, downregulation of expression of Igf2 and RasGrf1 together with upregulation of Igf2R, which is a decoy receptor for IGF2, keeps VSELs quiescent. At the same time, these cells also have high expression of H19 and p57Kip2, which are also regulated by imprinting, and negatively affect cell proliferation. However, with increasing age, the methylation pattern of DMRs at paternally imprinted genes in VSELs reverses andIgf2-H19, as well as RasGrf1, loci become gradually methylated, and thus VSELs become more sensitive to IIS [83], which may contribute to their age-related depletion over time.
On the other hand (Figure 2), VSELs express functional IGF-1R and InsR and are susceptible to exogenous circulating paracrine IGF1 and insulin (Ins). Therefore, a chronic increase in caloric uptake that elevates circulating levels of IGF1 and Ins may contribute over time to depletion of these cells from adult tissues, affect the generation of VSEL-derived TCSCs, and thus negatively affect life span. This explains why mice that have high levels of circulating blood plasma IGF1 and enhanced IIS display accelerated depletion of VSELs and have a shorter lifespan than age-matched littermates [85].
Conclusions
New data from our group and other groups has provided more evidence on the existence and biological role of primitive embryonic-like stem cells in murine adult tissues and their potential role in i) tissue organ rejuvenation, ii) longevity, and iii) regeneration/repair of damaged tissues.
While murine BM-derived VSELs have been extensively characterized, we are aware that more work is needed to better characterize these small cells at the molecular level in humans. We need to determine whether human VSELs have the same molecular signature (e.g., an open chromatin structure at the Oct4 promoter, modification of somatic imprinting, and the presence of bivalent domains) as their murine counterparts. If we can confirm that a similar mechanism operates for human and murine BM-derived UCB-VSELs, perhaps a controlled modulation of the somatic imprinted state to produce proper de novo methylation of somatic imprinted genes on the maternal and paternal chromosomes could increase the regenerative power of these cells.
Finally, our work on murine VSELs has for the first time connected the role of caloric restriction/uptake, IIS, and the number of VSELs playing a potential role in tissue and organ rejuvenation [83,85]. An increase in chronic caloric uptake leads to an increase in IIS and may prematurely deplete VSELs from adult tissues, thus contributing to aging. By contrast, caloric restriction has an opposing beneficial effect on the tissue-residing pool of VSELs and longevity. Based on the encouraging studies on VSELs in the murine BM, further studies are needed to evaluate whether the number of VSELs in other adult murine tissues is also sensitive to IIS and correlates with life span. Similar studies have also to be performed on human VSELs.
Acknowledgments
This work is supported by NIH grant R01 DK074720, the EU Innovative Economy Operational Program POIG.01.01.02-00-109/09-01, and the Stella and Henry Hoenig Endowment to MZR.
Conflicts of Interest
The authors of this manuscript have no conflict of interest to declare.
References
- 1. Ratajczak MZ, Zuba-Surma EK, Wysoczynski M, Wan W, Ratajczak J, Wojakowski W, Kucia M. Hunt for pluripotent stem cell - Regenerative medicine search for almighty cell. J Autoimmun. 2008; 30: 151 -162. [PubMed] .
- 2. D'Ippolito G, Diabira S, Howard GA, Menei P, Roos BA, Schiller PC. Marrow-isolated adult multilineage inducible (MIAMI) cells, a unique population of postnatal young and old human cells with extensive expansion and differentiation potential. J Cell Sci. 2004; 117: 2971 -2981. [PubMed] .
- 3. Asahara T, Murohara T, Sullivan A, Silver M, van der Zee R, Li T, Witzenbichler B, Schatteman G, Isner JM. Isolation of putative progenitor endothelial cells for angiogenesis. Science. 1997; 275: 964 -967. [PubMed] .
- 4. Kogler G, Sensken S, Airey JA, Trapp T, Muschen M, Feldhahn N, Liedtke S, Sorg RV, Fischer J, Rosenbaum C, et al. A new human somatic stem cell from placental cord blood with intrinsic pluripotent differentiation potential. J Exp Med. 2004; 200: 123 -135. [PubMed] .
- 5. Prockop DJ. Marrow stromal cells as stem cells for nonhematopoietic tissues. Science. 1997; 276: 71 -74. [PubMed] .
- 6. Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S, Marshak DR. Multilineage potential of adult human mesenchymal stem cells. Science. 1999; 284: 143 -147. [PubMed] .
- 7. Brill S, Holst P, Sigal S, Zvibel I, Fiorino A, Ochs A, Somasundaran U, Reid LM. Hepatic progenitor populations in embryonic, neonatal, and adult liver. Proc Soc Exp Biol Med. 1993; 204: 261 -269. [PubMed] .
- 8. Thomas DB. The differentiation of transplanted haematopoietic cells derived from bone marrow, spleen and fetal liver. J Anat. 1971; 110: 297 -306. [PubMed] .
- 9. Blanpain C, Horsley V, Fuchs E. Epithelial stem cells: turning over new leaves. Cell. 2007; 128: 445 -458. [PubMed] .
- 10. Wognum AW, Eaves AC, Thomas TE. Identification and isolation of hematopoietic stem cells. Arch Med Res. 2003; 34: 461 -475. [PubMed] .
- 11. Ratajczak MZ. Phenotypic and functional characterization of hematopoietic stem cells. Curr Opin Hematol. 2008; 15: 293 -300. [PubMed] .
- 12. Kucia M, Ratajczak J, Reca R, Janowska-Wieczorek A, Ratajczak MZ. Tissue-specific muscle, neural and liver stem/progenitor cells reside in the bone marrow, respond to an SDF-1 gradient and are mobilized into peripheral blood during stress and tissue injury. Blood Cells Mol Dis. 2004; 32: 52 -57. [PubMed] .
- 13. Ratajczak MZ, Kucia M, Reca R, Majka M, Janowska-Wieczorek A, Ratajczak J. Stem cell plasticity revisited: CXCR4-positive cells expressing mRNA for early muscle, liver and neural cells ‘hide out’ in the bone marrow. Leukemia. 2004; 18: 29 -40. [PubMed] .
- 14. Ratajczak MZ, Liu R, Marlicz W, Blogowski W, Starzynska T, Wojakowski W, Zuba-Surma E. Identification of very small embryonic/epiblast-like stem cells (VSELs) circulating in peripheral blood during organ/tissue injuries. Methods Cell Biol. 2011; 103: 31 -54. [PubMed] .
- 15. Ratajczak MZ, Liu R, Ratajczak J, Kucia M, Shin DM. The role of pluripotent embryonic-like stem cells residing in adult tissues in regeneration and longevity. Differentiation. 2011; 81: 153 -161. [PubMed] .
- 16. Zuba-Surma EK, Kucia M, Ratajczak J, Ratajczak MZ. “Small stem cells” in adult tissues: very small embryonic-like stem cells stand up!. Cytometry A. 2009; 75: 4 -13. [PubMed] .
- 17. McGuckin CP, Forraz N, Baradez MO, Navran S, Zhao J, Urban R, Tilton R, Denner L. Production of stem cells with embryonic characteristics from human umbilical cord blood. Cell Prolif. 2005; 38: 245 -255. [PubMed] .
- 18. Jiang Y, Jahagirdar BN, Reinhardt RL, Schwartz RE, Keene CD, Ortiz-Gonzalez XR, Reyes M, Lenvik T, Lund T, Blackstad M, et al. Pluripotency of mesenchymal stem cells derived from adult marrow. Nature. 2002; 418: 41 -49. [PubMed] .
- 19. Beltrami AP, Cesselli D, Bergamin N, Marcon P, Rigo S, Puppato E, D'Aurizio F, Verardo R, Piazza S, Pignatelli A, et al. Multipotent cells can be generated in vitro from several adult human organs (heart, liver and bone marrow). Blood. 2007; 110: 3438 -3446. [PubMed] .
- 20. Tondreau T, Meuleman N, Delforge A, Dejeneffe M, Leroy R, Massy M, Mortier C, Bron D, Lagneaux L. Mesenchymal stem cells derived from CD133-positive cells in mobilized peripheral blood and cord blood: proliferation, Oct4 expression, and plasticity. Stem Cells. 2005; 23: 1105 -12. [PubMed] .
- 21. Wernet P, Trapp T, Zweigerdt R, Mann J, Trompeter HI. Lentiviral labeling reveals three germ layer differentiation potential of a single unrestricted somatic stem cell from human cord blood. Exp Hematol. 2010; 38: 1099 -104. [PubMed] .
- 22. Santourlidis S, Wernet P, Ghanjati F, Graffmann N, Springer J, Kriegs C, Zhao X, Brands J, Arauzo-Bravo MJ, Neves R, et al. Unrestricted somatic stem cells (USSC) from human umbilical cord blood display uncommitted epigenetic signatures of the major stem cell pluripotency genes. Stem Cell Res. 2011; 6: 60 -69. [PubMed] .
- 23. Jones RJ, Collector MI, Barber JP, Vala MS, Fackler MJ, May WS, Griffin CA, Hawkins AL, Zehnbauer BA, Hilton J, et al. Characterization of mouse lymphohematopoietic stem cells lacking spleen colony-forming activity. Blood. 1996; 88: 487 -491. [PubMed] .
- 24. Jones RJ, Wagner JE, Celano P, Zicha MS, Sharkis SJ. Separation of pluripotent haematopoietic stem cells from spleen colony-forming cells. Nature. 1990; 347: 188 -189. [PubMed] .
- 25. Krause DS. Bone marrow-derived cells and stem cells in lung repair. Proc Am Thorac Soc. 2008; 5: 323 -327. [PubMed] .
- 26. Vacanti MP, Roy A, Cortiella J, Bonassar L, Vacanti CA. Identification and initial characterization of spore-like cells in adult mammals. J Cell Biochem. 2001; 80: 455 -460. [PubMed] .
- 27. Orlic D, Kajstura J, Chimenti S, Jakoniuk I, Anderson SM, Li B, Pickel J, McKay R, Nadal-Ginard B, Bodine DM, et al. Bone marrow cells regenerate infarcted myocardium. Nature. 2001; 410: 701 -705. [PubMed] .
- 28. Mezey E, Chandross KJ, Harta G, Maki RA, McKercher SR. Turning blood into brain: cells bearing neuronal antigens generat-ed in vivo from bone marrow. Science. 2000; 290: 1779 -1782. [PubMed] .
- 29. Orkin SH and Zon LI. Hematopoiesis and stem cells: plasticity versus developmental heterogeneity. Nat Immunol. 2002; 3: 323 -328. [PubMed] .
- 30. Wagers AJ, Sherwood RI, Christensen JL, Weissman IL. Little evidence for developmental plasticity of adult hematopoietic stem cells. Science. 2002; 297: 2256 -2259. [PubMed] .
- 31. Kucia M, Reca R, Campbell FR, Zuba-Surma E, Majka M, Ratajczak J, Ratajczak MZ. A population of very small embryonic-like (VSEL) CXCR4(+)SSEA-1(+)Oct-4+ stem cells identified in adult bone marrow. Leukemia. 2006; 20: 857 -869. [PubMed] .
- 32. Zuba-Surma EK, Kucia M, Abdel-Latif A, Dawn B, Hall B, Singh R, Lillard JW Jr., Ratajczak MZ. Morphological characterization of very small embryonic-like stem cells (VSELs) by ImageStream system analysis. J Cell Mol Med. 2008; 12: 292 -303. [PubMed] .
- 33. Zuba-Surma EK, Kucia M, Rui L, Shin DM, Wojakowski W, Ratajczak J, Ratajczak MZ. Fetal liver very small embryonic/epiblast like stem cells follow developmental migratory pathway of hematopoietic stem cells. Ann N Y Acad Sci. 2009; 1176: 205 -218. [PubMed] .
- 34. Zuba-Surma EK, Kucia M, Wu W, Klich I, Lillard JW Jr., Ratajczak J, Ratajczak MZ. Very small embryonic-like stem cells are present in adult murine organs: ImageStream-based morphological analysis and distribution studies. Cytometry A. 2008; 73A: 1116 -1127. [PubMed] .
- 35. Ratajczak MZ, Kucia M, Ratajczak J, Zuba-Surma EK. A multi-instrumental approach to identify and purify very small embryonic like stem cells (VSELs) from adult tissues. Micron. 2009; 40: 386 -393. [PubMed] .
- 36. Liu Y, Gao L, Zuba-Surma EK, Peng X, Kucia M, Ratajczak MZ, Wang W, Enzmann V, Kaplan HJ, Dean DC. Identification of small Sca-1(+), Lin(-), CD45(-) multipotential cells in the neonatal murine retina. Exp Hematol. 2009; 37: 1096 -107. 1107 e1 [PubMed] .
- 37. Kucia M, Halasa M, Wysoczynski M, Baskiewicz-Masiuk M, Moldenhawer S, Zuba-Surma E, Czajka R, Wojakowski W, Machalinski B, Ratajczak MZ. Morphological and molecular characterization of novel population of CXCR4(+) SSEA-4(+) Oct-4(+) very small embryonic-like cells purified from human cord blood - preliminary report. Leukemia. 2007; 21: 297 -303. [PubMed] .
- 38. Ratajczak J, Zuba-Surma E, Klich I, Liu R, Wysoczynski M, Greco N, Kucia M, Laughlin MJ, Ratajczak MZ. Hematopoietic differentiation of umbilical cord blood-derived very small embryonic/epiblast-like stem cells. Leukemia. 2011; 25: 1278 -1285. [PubMed] .
- 39. Zuba-Surma EK, Klich I, Greco N, Laughlin MJ, Ratajczak J, Ratajczak MZ. Optimization of isolation and further characterization of umbilical-cord-blood-derived very small embryonic/ epiblast-like stem cells (VSELs). Eur J Haematol. 2010; 84: 34 -46. [PubMed] .
- 40. Abdel-Latif A, Zuba-Surma EK, Ziada KM, Kucia M, Cohen DA, Kaplan AM, Van Zant G, Selim S, Smyth SS, Ratajczak MZ. Evidence of Mobilization of Pluripotent Stem Cells into Peripheral Blood of Patients with Myocardial Ischemia. Exp Hematol. 2010; .
- 41. Paczkowska E, Kucia M, Koziarska D, Halasa M, Safranow K, Masiuk M, Karbicka A, Nowik M, Nowacki P, Ratajczak MZ, et al. Clinical evidence that very small embryonic-like stem cells are mobilized into peripheral blood in patients after stroke. Stroke. 2009; 40: 1237 -1244. [PubMed] .
- 42. Wojakowski W, Tendera M, Kucia M, Zuba-Surma E, Paczkowska E, Ciosek J, Halasa M, Krol M, Kazmierski M, Buszman P, et al. Mobilization of bone marrow-derived Oct-4+ SSEA-4+ very small embryonic-like stem cells in patients with acute myocardial infarction. J Am Coll Cardiol. 2009; 53: 1 -9. [PubMed] .
- 43. Sovalat H, Scrofani M, Eidenschenk A, Pasquet S, Rimelen V, Henon P. Identification and isolation from either adult human bone marrow or G-CSF-mobilized peripheral blood of CD34(+)/CD133(+)/CXCR4(+)/ Lin(-)CD45(-) cells, featuring morphological, molecular, and phenotypic characteristics of very small embryonic-like (VSEL) stem cells. Exp Hematol. 2011; 39: 495 -505. [PubMed] .
- 44. Zuba-Surma EK and Ratajczak MZ. Overview of very small embryonic-like stem cells (VSELs) and methodology of their identification and isolation by flow cytometric methods. Curr Protoc Cytom. 2010; Chapter 9:Unit9 29 .
- 45. Iskovich S, Goldenberg-Cohen N, Stein J, Yaniv I, Fabian I, Askenasy N. Elutriated stem cells derived from the adult bone marrow differentiate into insulin-producing cells in vivo and reverse chemical diabetes. Stem Cells Dev. 2012; 21: 86 -96. [PubMed] .
- 46. Kassmer SH, Bruscia EM, Zhang PX, Krause DS. Non-Hematopoietic Cells are the Primary Source of Bone Marrow Derived Lung Epithelial Cells. Stem Cells. 2011; .
- 47. Virant-Klun I and Skutella T. Stem cells in aged mammalian ovaries. Aging (Albany NY). 2010; 2: 3 -6. [PubMed] .
- 48. Kuroda Y, Kitada M, Wakao S, Nishikawa K, Tanimura Y, Makinoshima H, Goda M, Akashi H, Inutsuka A, Niwa A, et al. Unique multipotent cells in adult human mesenchymal cell populations. Proc Natl Acad Sci U S A. 2010; 107: 8639 -8643. [PubMed] .
- 49. Taichman RS, Wang Z, Shiozawa Y, Jung Y, Song J, Balduino A, Wang J, Patel LR, Havens AM, Kucia M, et al. Prospective identification and skeletal localization of cells capable of multilineage differentiation in vivo. Stem Cells Dev. 2010; 19: 1557 -1570. [PubMed] .
- 50. Parte S, Bhartiya D, Telang J, Daithankar V, Salvi V, Zaveri K, Hinduja I. Detection, characterization, and spontaneous differentiation in vitro of very small embryonic-like putative stem cells in adult Mammalian ovary. Stem Cells Dev. 2011; 20: 1451 -1464. [PubMed] .
- 51. Bhartiya D, Kasiviswanathan S, Unni SK, Pethe P, Dhabalia JV, Patwardhan S, Tongaonkar HB. Newer insights into premeiotic development of germ cells in adult human testis using Oct-4 as a stem cell marker. J Histochem Cytochem. 2010; 58: 1093 -1106. [PubMed] .
- 52. Wu JH, Wang HJ, Tan YZ, Li ZH. Characterization of Rat Very Small Embryonic-Like Stem Cells and Cardiac Repair After Cell Transplantation for Myocardial Infarction. Stem Cells Dev. 2011; .
- 53. Goldenberg-Cohen N, Avraham-Lubin BC, Sadikov T, Goldstein RS, Askenasy N. Primitive Stem Cells Derived from Bone Marrow Express Glial and Neuronal Markers and Support Revascularization in Injured Retina Exposed to Ischemic and Mechanical Damage. Stem Cells Dev. 2011; .
- 54. Wakao S, Kitada M, Kuroda Y, Shigemoto T, Matsuse D, Akashi H, Tanimura Y, Tsuchiyama K, Kikuchi T, Goda M, et al. Multilineage-differentiating stress-enduring (Muse) cells are a primary source of induced pluripotent stem cells in human fibroblasts. Proc Natl Acad Sci U S A. 2011; 108: 9875 -9880. [PubMed] .
- 55. Bhartiya D, Shaikh A, Nagvenkar P, Kasiviswanathan S, Pethe P, Pawani H, Mohanty S, Rao SG, Zaveri K, Hinduja I. Very small embryonic-like stem cells with maximum regenerative potential get discarded during cord blood banking and bone marrow processing for autologous stem cell therapy. Stem Cells Dev. 2012; 21: 1 -6. [PubMed] .
- 56. McGuckin C, Jurga M, Ali H, Strbad M, Forraz N. Culture of embryonic-like stem cells from human umbilical cord blood and onward differentiation to neural cells in vitro. Nat Protoc. 2008; 3: 1046 -1055. [PubMed] .
- 57. Seo KW, Lee SR, Bhandari DR, Roh KH, Park SB, So AY, Jung JW, Seo MS, Kang SK, Lee YS, et al. OCT4A contributes to the stemness and multi-potency of human umbilical cord blood-derived multipotent stem cells (hUCB-MSCs). Biochem Biophys Res Commun. 2009; 384: 120 -125. [PubMed] .
- 58. Gordon MY, Levicar N, Pai M, Bachellier P, Dimarakis I, Al-Allaf F, M'Hamdi H, Thalji T, Welsh JP, Marley SB, et al. Characterization and clinical application of human CD34+ stem/progenitor cell populations mobilized into the blood by granulocyte colony-stimulating factor. Stem Cells. 2006; 24: 1822 -1830. [PubMed] .
- 59. Mikhail MA, M'Hamdi H, Welsh J, Levicar N, Marley SB, Nicholls JP, Habib NA, Louis LS, Fisk NM, Gordon MY. High frequency of fetal cells within a primitive stem cell population in maternal blood. Hum Reprod. 2008; 23: 928 -933. [PubMed] .
- 60. Moreb JS. Aldehyde dehydrogenase as a marker for stem cells. Curr Stem Cell Res Ther. 2008; 3: 237 -246. [PubMed] .
- 61. Hess DA, Wirthlin L, Craft TP, Herrbrich PE, Hohm SA, Lahey R, Eades WC, Creer MH, Nolta JA. Selection based on CD133 and high aldehyde dehydrogenase activity isolates long-term reconstituting human hematopoietic stem cells. Blood. 2006; 107: 2162 -2169. [PubMed] .
- 62. Wojakowski W, Tendera M, Michalowska A, Majka M, Kucia M, Maslankiewicz K, Wyderka R, Ochala A, Ratajczak MZ. Mobilization of CD34/CXCR4+, CD34/CD117+, c-met+ stem cells, and mononuclear cells expressing early cardiac, muscle, and endothelial markers into peripheral blood in patients with acute myocardial infarction. Circulation. 2004; 110: 3213 -3220. [PubMed] .
- 63. Drukala J, Paczkowska E, Kucia M, Mlynska E, Krajewski A, Machalinski B, Madeja Z, Ratajczak MZ. Stem Cells, Including a Population of Very Small Embryonic-Like Stem Cells, are Mobilized Into Peripheral Blood in Patients After Skin Burn Injury. Stem Cell Rev. 2011; .
- 64. Marlicz W, Zuba-Surma E, Kucia M, Blogowski W, Starzynska T, Ratajczak MZ. Various types of stem cells, including a population of very small embryonic-like stem cells, are mobilized into peripheral blood in patients with Crohn's disease. Inflamm Bowel Dis. 2012; .
- 65. Shin DM, Liu R, Klich I, Ratajczak J, Kucia M, Ratajczak MZ. Molecular characterization of isolated from murine adult tissues very small embryonic/epiblast like stem cells (VSELs). Mol Cells. 2010; 29: 533 -538. [PubMed] .
- 66. Shin DM, Liu R, Klich I, Wu W, Ratajczak J, Kucia M, Ratajczak MZ. Molecular signature of adult bone marrow-purified very small embryonic-like stem cells supports their developmental epiblast/germ line origin. Leukemia. 2010; 24: 1450 -1461. [PubMed] .
- 67. Shin DM, Zub-Surma EK, Ratajczak MZ, Kucia M. The Unique Pattern of Somatic Imprint in Oct-4+ Very Small Embryonic Like (VSEL) Stem Cells Isolated from Adult Tissues Further Supports Both Their Epiblast/Germ Line Origin and Explains Quiescent Status: Potential Modification of Somatic Imprint as a Key to Longevity? Blood. 2008; 112: 385 .
- 68. Shin DM, Liu R, Wu W, Waigel SJ, Zacharias W, Ratajczak MZ, Kucia M. Global Gene Expression Analysis of Very Small Embryonic-Like Stem Cells Reveals that the Ezh2-Dependent Bivalent Domain Mechanism Contributes to Their Pluripotent State. Stem Cells Dev. 2011; .
- 69. Muller FJ, Goldmann J, Loser P, Loring JF. A call to standardize teratoma assays used to define human pluripotent cell lines. Cell Stem Cell. 2010; 6: 412 -414. [PubMed] .
- 70. Smith KP, Luong MX, Stein GS. Pluripotency: toward a gold standard for human ES and iPS cells. J Cell Physiol. 2009; 220: 21 -29. [PubMed] .
- 71. Zuba-Surma EK, Kucia M, Dawn B, Guo Y, Ratajczak MZ, Bolli R. Bone marrow-derived pluripotent very small embryonic-like stem cells (VSELs) are mobilized after acute myocardial infarction. J Mol Cell Cardiol. 2008; 44: 865 -873. [PubMed] .
- 72. Shin DM, Zuba-Surma EK, Wu W, Ratajczak J, Wysoczynski M, Ratajczak MZ, Kucia M. Novel epigenetic mechanisms that control pluripotency and quiescence of adult bone marrow-derived Oct4(+) very small embryonic-like stem cells. Leukemia. 2009; 23: 2042 -2051. [PubMed] .
- 73. Ratajczak J, Wysoczynski M, Zuba-Surma E, Wan W, Kucia M, Yoder MC, Ratajczak MZ. Adult murine bone marrow-derived very small embryonic-like stem cells differentiate into the hematopoietic lineage after coculture over OP9 stromal cells. Exp Hematol. 2011; 39: 225 -237. [PubMed] .
- 74. Dawn B, Tiwari S, Kucia MJ, Zuba-Surma EK, Guo Y, Sanganalmath SK, Abdel-Latif A, Hunt G, Vincent RJ, Taher H, et al. Transplantation of bone marrow-derived very small embryonic-like stem cells attenuates left ventricular dysfunction and remodeling after myocardial infarction. Stem Cells. 2008; 26: 1646 -1655. [PubMed] .
- 75. Zuba-Surma EK, Guo Y, Taher H, Sanganalmath SK, Hunt G, Vincent RJ, Kucia M, Abdel-Latif A, Tang XL, Ratajczak MZ, et al. Transplantation of expanded bone marrow-derived very small embryonic-like stem cells (VSEL-SCs) improves left ventricular function and remodelling after myocardial infarction. J Cell Mol Med. 2011; 15: 1319 -1328. [PubMed] .
- 76. Ratajczak MZ, Zuba-Surma EK, Shin DM, Ratajczak J, Kucia M. Very small embryonic-like (VSEL) stem cells in adult organs and their potential role in rejuvenation of tissues and longevity. Exp Gerontol. 2008; .
- 77. Zuba-Surma EK and Ratajczak MZ. Very Small Embryonic Like Stem Cells - implications for aging. Mech Ageing Dev. .
- 78. Reik W and Walter J. Genomic imprinting: parental influence on the genome. Nat Rev Genet. 2001; 2: 21 -32. [PubMed] .
- 79. Yamazaki Y, Mann MR, Lee SS, Marh J, McCarrey JR, Yanagimachi R, Bartolomei MS. Reprogramming of primordial germ cells begins before migration into the genital ridge, making these cells inadequate donors for reproductive cloning. Proc Natl Acad Sci U S A. 2003; 100: 12207 -12212. [PubMed] .
- 80. Wu Q, Kumagai T, Kawahara M, Ogawa H, Hiura H, Obata Y, Takano R, Kono T. Regulated expression of two sets of paternally imprinted genes is necessary for mouse parthenogenetic development to term. Reproduction. 2006; 131: 481 -488. [PubMed] .
- 81. Park CH, Uh KJ, Mulligan BP, Jeung EB, Hyun SH, Shin T, Ka H, Lee CK. Analysis of imprinted gene expression in normal fertilized and uniparental preimplantation porcine embryos. PLoS One. 2011; 6: e22216 [PubMed] .
- 82. Ratajczak MZ, Shin DM, Ratajczak J, Kucia M, Bartke A. A novel insight into aging: are there pluripotent very small embryonic-like stem cells (VSELs) in adult tissues overtime depleted in an Igf-1-dependent manner? Aging (Albany NY). 2010; 2: 875 -883. [PubMed] .
- 83. Kucia M, Masternak M, Liu R, Shin DM, Ratajczak J, Mierzejewska K, Spong A, Kopchick JJ, Bartke A, Ratajczak MZ. The negative effect of prolonged somatotrophic/insulin signaling on an adult bone marrow-residing population of pluripotent very small embryonic-like stem cells (VSELs). Age (Dordr). 2012; .
- 84. Svensson J, Sjogren K, Faldt J, Andersson N, Isaksson O, Jansson JO, Ohlsson C. Liver-derived IGF-I regulates mean life span in mice. PLoS One. 2011; 6: e22640 [PubMed] .
- 85. Kucia M, Shin DM, Liu R, Ratajczak J, Bryndza E, Masternak MM, Bartke A, Ratajczak MZ. Reduced number of VSELs in the bone marrow of growth hormone transgenic mice indicates that chronically elevated Igf1 level accelerates age-dependent exhaustion of pluripotent stem cell pool: a novel view on aging. Leukemia. 2011; 25: 1370 -1374. [PubMed] .