



Introduction
Hormesis (a neologism coined from the ancient Greek term hormáein, which literally means “to set in motion, impel, urge on”) describes a favorable biological response to harmless doses of toxins and other stressors. Hormesis-stimulating compounds initiate an adaptive stress response that renders cells/organisms resistant against high (and normally harmful) doses of the same agent. On the theoretical level, hormesis may constitute (one of) the mechanisms that allows stressed cells to avoid senescence and death, and hence might have some impact on the (patho)physiology of aging. Thus, measures that reportedly prolong the healthy lifespan of multiple species, such as caloric restriction and the administration of resveratrol [1-6], may do so by inducing a hormetic response [7,8]. In this article, we will examine the molecular circuitries that link cellular stress and death, and how these pathways can get uncoupled during hormetic responses.
Redundant pathways leading to apoptotic cell death
Apoptosis is frequently viewed as a caspase-dependent cell death pathway in which a series of specific cysteine proteases are activated in a cascade of proteolytic maturation steps [9,10]. In response to cell death-inducing signals, so-called initiator caspases (i.e., caspase-8 and -9) [11,12] get engaged and activate so-called effector caspases (i.e., caspase-3, -6 and -7) [13], which in turn degrade multiple proteins causing the arrest of vital cellular functions as well as the initiation of lethal catabolic reactions [14-16]. Two upstream events account for the activation of initiator caspases. In the extrinsic pathway, caspase-8 is recruited to and gets activated within the death-inducing signaling complex (DISC), a multiprotein complex that forms at the cytoplasmic tails of a specific class of cell surface receptors, the death receptors, upon their occupancy by their respective ligands [12,17-19]. In the intrinsic pathway, caspase-9 is activated at the so-called apoptosome, a supramolecular entity that involves dATP, the cytoplasmic protein APAF1 and the mitochondrial intermembrane space factor cytochrome c, and that only forms when the outer mitochondrial membrane, which usually separates APAF1 (outside) and cytochrome c (inside) is permeabilized [20-24].
Nonetheless, caspase inhibition rarely prevents cell death completely (although it sometimes attenuates the morphological manifestations of apoptosis) [10,25-27], and multiple caspase-independent pathways may come into action [9,28,29]. For example, permeabilized mitochondria can allow for the release of apoptosis-inducing factor (AIF) and cofilin, both of which have been shown to operate as caspase-independent death effectors [30-33]. This places the control of cell death at the level of the mitochondrion, and more precisely at the level of mitochondrial membranes, whose permeabilization is controlled by multiple upstream effectors and processes, including distinct classes of stress-activated kinases [24,34-43], the tumor suppressor protein p53 [44-46], epigenetic perturbations [47-51], (de)acetylases [52,53], perturbations of the cell cycle [54,55], and nuclear damage [56-60].
In accord with the key role of mitochondria in the regulation of many (if not all) apoptotic pathways [61-65], mitochondrial functions and integrity are controlled by a variety of distinct mechanisms. Proteins from the Bcl-2 family are considered as central modulators of mitochondrial apoptosis [66-68], but other proteins that are not directly related to BCL-2 can induce or suppress mitochondrial outer membrane permeabilization (MOMP) as well. Thus, beyond BCL2 and its close relatives BCL2L1 (best known as Bcl-XL) and MCL1, PRELI [69], the uncoupling protein 2 (UCP2) [70] and the X-linked inhibitor of apoptosis protein (XIAP) [22] can prevent MOMP. Pro-apoptotic proteins like the multidomain proteins BAX and BAK [71], as well as multiple BH3-only proteins stimulate MOMP [72-74]. In addition, pro-oxidants [75-77], membrane-destabilizing lipids (such as ceramide) [52] and free Ca2+ ions (modulated by other divalent cations such as Mg2+ and Zn2+, and possibly by the monovalent cation Li+) [21,23,78-80] can stimulate MOMP.
Autophagy as a cytoprotective mechanism
Macroautophagy (to which we refer to as “autophagy”) is a lysosomal degradation pathway in which portions of the cytoplasm (organelles or cytosol) are enwrapped in double-membraned vesicles (called autophagosomes) that fuse with lysosomes and get degraded by lysosomal hydrolases [81-84]. Importantly, autophagy and apoptosis exhibit a consistent degree of crosstalk, at multiple levels [85-87].
Autophagy can lead to the removal of damaged, potentially dangerous mitochondria, thereby increasing the threshold for cell death induction by MOMP-inducing agents or other stressors. Thus, both mitochondrion-specific autophagy (mitophagy) and general autophagy can reduce the propensity of cells to undergo apoptosis [1,76,88-90].
Caspase-dependent apoptosis is associated with the degradation of Beclin 1 by caspases. As Beclin 1 is essential for the initial steps of autophagy, caspase activation most often result into the inhibition of the autophagic pathway [16,91,92]. This reflects a general pattern according to which pro-apoptotic signals result in the inhibition of pro-survival systems.
Some molecular mechanisms that sense cellular stress can induce both autophagy and apoptosis. This applies for instance to BH3 proteins (as well as pharmacological BH3 mimetics), which can liberate Beclin 1 from inhibitory interactions with BCL-2-like proteins, thereby favoring autophagy, and also stimulate MOMP by activating BAX or BAK [93-101]. It is thought that the relative abundance of different Bcl-2 family members, as well as their subcellular localization and activation state may determine whether BH3 proteins/mimetics induce autophagy or apoptosis [66,67]. Moreover, endoplasmic reticulum stress can either result in autophagy or in apoptosis, depending on a yet-to-elucidated interplay among threshold effects [88,102,103]. One possible scenario suggests that mild stress would induce an autophagic response that elevates the threshold for apoptosis induction. This would represent a typical case of hormesis (Figure 1).
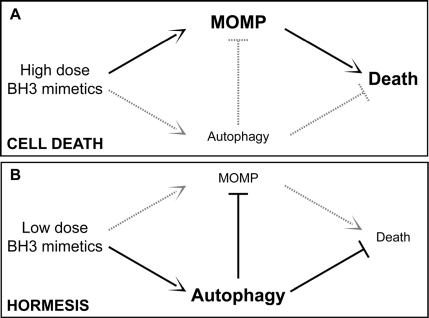
Figure 1. Autophagy and hormesis In baseline conditions, autophagy contributes to the maintenance of cellular homeostasis by removing potentially dangerous mitochondria (or other damaged organelles) and by ensuring the disposal of protein aggregates. This might have anti-aging effects and prolong healthy lifespan by elevating the threshold of damage required for the induction of cellular dysfunctions or death. In this scenario, while high doses of agents that stimulate both autophagy and cell death (e.g., BH3 mimetics) would be toxic (A), low doses of the same agents might provoke a hormetic response and hence favor the adaptation of cells to stress (B). MOMP, mitochondrial outer membrane permeabilization.
Autophagy as an anti-aging mechanism
Pharmacological or genetic manipulations designed to prolong lifespan induce autophagy in multiple model organisms, including yeast, nematodes and flies, and the inhibition of autophagy often (always?) prevents longevity extension in such settings. This applies to lifespan extension induced by caloric restriction, genetic or pharmacological activation of Sirtuin 1, inhibition of the mammalian target of rapamycin (mTOR) with rapamycin, and administration of spermidine, a histone acetylase inhibitor [3,4,81,104-107]. Among these stimuli, there is circumstantial evidence that Sirtuin 1 (whose activation occurs during and is necessary for starvation- and resveratrol-induced autophagy) acts in a hormetic fashion. One of the best-known systems of hormesis is ischemic preconditioning (IPC), whereby short episodes of ischemia protect the brain against a later, more severe reduction in oxygen and nutrient supply. In this system, the administration of resveratrol can mimic IPC, and both resveratrol and IPC induce similar changes in the acetylproteome of the brain [53].
It is currently a mystery through which mechanisms autophagy may increase lifespan [108,109]. One obvious possibility resides in the cytoprotective, apoptosis-inhibitory action of autophagy (see above), although other possibilities must be considered as well. For example, there is ample evidence that the inhibition of mTOR (which stimulates autophagy, yet may have other metabolic effects as well) antagonizes senescence [105,110-115]. Thus, it can be speculated, yet remains to be proven, that autophagy would avoid cellular senescence induced by DNA-damaging agents [116-119], as well as the attrition of stem cells that accompanies advanced aging [120-123]. Simultaneously, there is strong evidence indicating that autophagy functions as an onco-suppressive mechanism that avoids the genetic instability that accelerates multi-step oncogenesis [3,114,124-130].
In addition, autophagy enhances the turnover of aggregate-prone cellular proteins, thus reducing the abundance of potent neurotoxic factors including, but not limited to, huntingtin aggregates [131]. Whether the induction of autophagy may also affect the accumulation of potentially toxic extracellular proteins (such as amyloid β and others) remains to be clarified [32,132-134].
At this stage, it is not clear which (if any) among these putative mechanisms plays a preponderant role in the longevity-increasing potential of autophagy. Future work will have to clarify this issue, which may have a major impact on how we design strategies for prolonging healthy lifespan.
Acknowledgments
GK is supported by the Ligue Nationale contre le Cancer (Equipe labellisé), Agence Nationale pour la Recherche (ANR), AXA Chair for Longevity Research, European Commission (Active p53, Apo-Sys, ChemoRes, ApopTrain), Fondation pour la Recherche Médicale (FRM), Institut National du Cancer (INCa), Cancéropôle Ile-de-France, Fondation Bettencourt-Schueller and the LabEx Onco-Immunology.
Conflicts of Interest
The authors of this manuscript have no conflict of interest to declare.
References
- 1. Blagosklonny MV. Linking calorie restriction to longevity through sirtuins and autophagy: any role for TOR. Cell Death Dis. 2010; 1 -e12. .
- 2. Fontana L, Partridge L, Longo VD. Extending healthy life span–from yeast to humans. Science. 2010; 328: 321 -6. [PubMed] .
- 3. Morselli E, Galluzzi L, Kepp O, Vicencio JM, Criollo A, Maiuri MC, Kroemer G. Anti- and pro-tumor functions of autophagy. Biochim Biophys Acta. 2009; 1793: 1524 -32. [PubMed] .
- 4. Morselli E, Maiuri MC, Markaki M, Megalou E, Pasparaki A, Palikaras K, Criollo A, Galluzzi L, Malik SA, Vitale I, Michaud M, Madeo F, Tavernarakis N, et al. Caloric restriction and resveratrol promote longevity through the Sirtuin-1-dependent induction of autophagy. Cell Death Dis. 2010; 1 -e10. .
- 5. Kirkland JL. Perspectives on cellular senescence and short term dietary restriction in adults. Aging (Albany NY). 2010; 2: 894 -6. [PubMed] .
- 6. Galikova M and Flatt T. Dietary restriction and other lifespan extending pathways converge at the activation of the downstream effector takeout. Aging (Albany NY). 2010; 2: 387 -9. [PubMed] .
- 7. Kouda K and Iki M. Beneficial effects of mild stress (hormetic effects): dietary restriction and health. J Physiol Anthropol. 2010; 29: 127 -32. [PubMed] .
- 8. Calabrese EJ, Mattson MP, Calabrese V. Resveratrol commonly displays hormesis: occurrence and biomedical significance. Hum Exp Toxicol. 2010; 29: 980 -1015. [PubMed] .
- 9. Galluzzi L, Vitale I, Abrams JM, Alnemri ES, Baehrecke EH, Blagosklonny MV, Dawson TM, Dawson VL, El-Deiry WS, Fulda S, Gottlieb E, Green DR, Hengartner MO, et al. Molecular definitions of cell death subroutines: recommendations of the Nomenclature Committee on Cell Death 2012. Cell Death Differ. 2011; in press. .
- 10. Kroemer G, Galluzzi L, Vandenabeele P, Abrams J, Alnemri ES, Baehrecke EH, Blagosklonny MV, El-Deiry WS, Golstein P, Green DR, Hengartner M, Knight RA, Kumar S, et al. Classification of cell death: recommendations of the Nomenclature Committee on Cell Death 2009. Cell Death Differ. 2009; 16: 3 -11. [PubMed] .
- 11. Janson V, Johansson A, Grankvist K. Resistance to caspase-8 and -9 fragments in a malignant pleural mesothelioma cell line with acquired cisplatin-resistance. Cell Death Dis. 2010; 1 -e78. .
- 12. Jalmar O, Garcia-Saez AJ, Berland L, Gonzalvez F, Petit PX. Giant unilamellar vesicles (GUVs) as a new tool for analysis of caspase-8/Bid-FL complex binding to cardiolipin and its functional activity. Cell Death Dis. 2010; 1 -e103. .
- 13. Sivananthan SN, Lee AW, Goodyer CG, LeBlanc AC. Familial amyloid precursor protein mutants cause caspase-6-dependent but amyloid beta-peptide-independent neuronal degeneration in primary human neuron cultures. Cell Death Dis. 2010; 1 -e100. .
- 14. Cheng JP, Betin VM, Weir H, Shelmani GM, Moss DK, Lane JD. Caspase cleavage of the Golgi stacking factor GRASP65 is required for Fas/CD95-mediated apoptosis. Cell Death Dis. 2010; 1 -e82. .
- 15. Tadokoro D, Takahama S, Shimizu K, Hayashi S, Endo Y, Sawasaki T. Characterization of a caspase-3-substrate kinome using an N- and C-terminally tagged protein kinase library produced by a cell-free system. Cell Death Dis. 2010; 1 -e89. .
- 16. Wirawan E, Vande Walle L, Kersse K, Cornelis S, Claerhout S, Vanoverberghe I, Roelandt R, De Rycke R, Verspurten J, Declercq W, Agostinis P, Vanden Berghe T, Lippens S, et al. Caspase-mediated cleavage of Beclin-1 inactivates Beclin-1-induced autophagy and enhances apoptosis by promoting the release of proapoptotic factors from mitochondria. Cell Death Dis. 2010; 1 -e18. .
- 17. Guardiola-Serrano F, Rossin A, Cahuzac N, Luckerath K, Melzer I, Mailfert S, Marguet D, Zornig M, Hueber AO. Palmitoylation of human FasL modulates its cell death-inducing function. Cell Death Dis. 2010; 1 -e88. .
- 18. Reis CR, van der Sloot AM, Natoni A, Szegezdi E, Setroikromo R, Meijer M, Sjollema K, Stricher F, Cool RH, Samali A, Serrano L, Quax WJ. Rapid and efficient cancer cell killing mediated by high-affinity death receptor homotrimerizing TRAIL variants. Cell Death Dis. 2010; 1 -e83. .
- 19. Schneider B, Munkel S, Krippner-Heidenreich A, Grunwald I, Wels WS, Wajant H, Pfizenmaier K, Gerspach J. Potent antitumoral activity of TRAIL through generation of tumor-targeted single-chain fusion proteins. Cell Death Dis. 2010; 1 -e68. .
- 20. Kroemer G, Galluzzi L, Brenner C. Mitochondrial membrane permeabilization in cell death. Physiol Rev. 2007; 87: 99 -163. [PubMed] .
- 21. Corona C, Masciopinto F, Silvestri E, Viscovo AD, Lattanzio R, Sorda RL, Ciavardelli D, Goglia F, Piantelli M, Canzoniero LM, Sensi SL. Dietary zinc supplementation of 3xTg-AD mice increases BDNF levels and prevents cognitive deficits as well as mitochondrial dysfunction. Cell Death Dis. 2010; 1 -e91. .
- 22. Flanagan L, Sebastia J, Tuffy LP, Spring A, Lichawska A, Devocelle M, Prehn JH, Rehm M. XIAP impairs Smac release from the mitochondria during apoptosis. Cell Death Dis. 2010; 1 -e49. .
- 23. Ruiz A, Matute C, Alberdi E. Intracellular Ca2+ release through ryanodine receptors contributes to AMPA receptor-mediated mitochondrial dysfunction and ER stress in oligodendrocytes. Cell Death Dis. 2010; 1 -e54. .
- 24. Tomiyama A, Tachibana K, Suzuki K, Seino S, Sunayama J, Matsuda KI, Sato A, Matsumoto Y, Nomiya T, Nemoto K, Yamashita H, Kayama T, Ando K, et al. MEK-ERK-dependent multiple caspase activation by mitochondrial proapoptotic Bcl-2 family proteins is essential for heavy ion irradiation-induced glioma cell death. Cell Death Dis. 2010; 1 -e60. .
- 25. Galluzzi L, Maiuri MC, Vitale I, Zischka H, Castedo M, Zitvogel L, Kroemer G. Cell death modalities: classification and pathophysiological implications. Cell Death Differ. 2007; 14: 1237 -43. [PubMed] .
- 26. Yuan J and Kroemer G. Alternative cell death mechanisms in development and beyond. Genes Dev. 2010; 24: 2592 -602. [PubMed] .
- 27. Cordeiro MF, Guo L, Coxon KM, Duggan J, Nizari S, Normando EM, Sensi SL, Sillito AM, Fitzke FW, Salt TE, Moss SE. Imaging multiple phases of neurodegeneration: a novel approach to assessing cell death in vivo. Cell Death Dis. 2010; 1 -e3. .
- 28. Vandenabeele P, Galluzzi L, Vanden Berghe T, Kroemer G. Molecular mechanisms of necroptosis: an ordered cellular explosion. Nat Rev Mol Cell Biol. 2010; 11: 700 -14. [PubMed] .
- 29. Wang Y, Nangia-Makker P, Balan V, Hogan V, Raz A. Calpain activation through galectin-3 inhibition sensitizes prostate cancer cells to cisplatin treatment. Cell Death Dis. 2010; 1 -e101. .
- 30. Patterson SD, Spahr CS, Daugas E, Susin SA, Irinopoulou T, Koehler C, Kroemer G. Mass spectrometric identification of proteins released from mitochondria undergoing permeability transition. Cell Death Differ. 2000; 7: 137 -44. [PubMed] .
- 31. Wabnitz GH, Goursot C, Jahraus B, Kirchgessner H, Hellwig A, Klemke M, Konstandin MH, Samstag Y. Mitochondrial translocation of oxidized cofilin induces caspase-independent necrotic-like programmed cell death of T cells. Cell Death Dis. 2010; 1 -e58. .
- 32. Lee MH, Lin SR, Chang JY, Schultz L, Heath J, Hsu LJ, Kuo YM, Hong Q, Chiang MF, Gong CX, Sze CI, Chang NS. TGF-beta induces TIAF1 self-aggregation via type II receptor-independent signaling that leads to generation of amyloid beta plaques in Alzheimer's disease. Cell Death Dis. 2010; 1 -e110. .
- 33. Osato K, Sato Y, Ochiishi T, Osato A, Zhu C, Sato M, Swanpalmer J, Modjtahedi N, Kroemer G, Kuhn HG, Blomgren K. Apoptosis-inducing factor deficiency decreases the proliferation rate and protects the subventricular zone against ionizing radiation. Cell Death Dis. 2010; 1 -e84. .
- 34. Allen-Petersen BL, Miller MR, Neville MC, Anderson SM, Nakayama KI, Reyland ME. Loss of protein kinase C delta alters mammary gland development and apoptosis. Cell Death Dis. 2010; 1 -e17. .
- 35. Chu KM, Minogue S, Hsuan JJ, Waugh MG. Differential effects of the phosphatidylinositol 4-kinases, PI4KIIalpha and PI4KIIIbeta, on Akt activation and apoptosis. Cell Death Dis. 2010; 1 -e106. .
- 36. Paoletti R, Maffei A, Madaro L, Notte A, Stanganello E, Cifelli G, Carullo P, Molinaro M, Lembo G, Bouche M. Protein kinase Ctheta is required for cardiomyocyte survival and cardiac remodeling. Cell Death Dis. 2010; 1 -e45. .
- 37. Jiang CC, Lai F, Tay KH, Croft A, Rizos H, Becker TM, Yang F, Liu H, Thorne RF, Hersey P, Zhang XD. Apoptosis of human melanoma cells induced by inhibition of B-RAFV600E involves preferential splicing of bimS. Cell Death Dis. 2010; 1 -e69. .
- 38. Fricker M, O'Prey J, Tolkovsky AM, Ryan KM. Phosphorylation of Puma modulates its apoptotic function by regulating protein stability. Cell Death Dis. 2010; 1 -e59. .
- 39. Pasupuleti N, Matsuyama S, Voss O, Doseff AI, Song K, Danielpour D, Nagaraj RH. The anti-apoptotic function of human alphaA-crystallin is directly related to its chaperone activity. Cell Death Dis. 2010; 1 -e31. .
- 40. Brandt B, Abou-Eladab EF, Tiedge M, Walzel H. Role of the JNK/c-Jun/AP-1 signaling pathway in galectin-1-induced T-cell death. Cell Death Dis. 2010; 1 -e23. .
- 41. Yuan M, Luong P, Hudson C, Gudmundsdottir K, Basu S. c-Abl phosphorylation of DeltaNp63alpha is critical for cell viability. Cell Death Dis. 2010; 1 -e16. .
- 42. Sears D, Luong P, Yuan M, Nteliopoulos G, Man YK, Melo JV, Basu S. Functional phosphoproteomic analysis reveals cold-shock domain protein A to be a Bcr-Abl effector-regulating proliferation and transformation in chronic myeloid leukemia. Cell Death Dis. 2010; 1 -e93. .
- 43. Ruela-de-Sousa RR, Fuhler GM, Blom N, Ferreira CV, Aoyama H, Peppelenbosch MP. Cytotoxicity of apigenin on leukemia cell lines: implications for prevention and therapy. Cell Death Dis. 2010; 1 -e19. .
- 44. Galluzzi L, Morselli E, Kepp O, Tajeddine N, Kroemer G. Targeting p53 to mitochondria for cancer therapy. Cell Cycle. 2008; 7: 1949 -55. [PubMed] .
- 45. Galluzzi L, Morselli E, Kepp O, Vitale I, Pinti M, Kroemer G. Mitochondrial liaisons of p53. Antioxid Redox Signal. 2011; 15: 1691 -714. [PubMed] .
- 46. Morselli E, Galluzzi L, Kroemer G. Mechanisms of p53-mediated mitochondrial membrane permeabilization. Cell Res. 2008; 18: 708 -10. [PubMed] .
- 47. Yelamanchili SV, Chaudhuri AD, Chen LN, Xiong H, Fox HS. MicroRNA-21 dysregulates the expression of MEF2C in neurons in monkey and human SIV/HIV neurological disease. Cell Death Dis. 2010; 1 -e77. .
- 48. Tenedini E, Roncaglia E, Ferrari F, Orlandi C, Bianchi E, Bicciato S, Tagliafico E, Ferrari S. Integrated analysis of microRNA and mRNA expression profiles in physiological myelopoiesis: role of hsa-mir-299-5p in CD34+ progenitor cells commitment. Cell Death Dis. 2010; 1 -e28. .
- 49. Sancho-Pelluz J, Alavi MV, Sahaboglu A, Kustermann S, Farinelli P, Azadi S, van Veen T, Romero FJ, Paquet-Durand F, Ekstrom P. Excessive HDAC activation is critical for neurodegeneration in the rd1 mouse. Cell Death Dis. 2010; 1 -e24. .
- 50. Lian J, Tian H, Liu L, Zhang XS, Li WQ, Deng YM, Yao GD, Yin MM, Sun F. Downregulation of microRNA-383 is associated with male infertility and promotes testicular embryonal carcinoma cell proliferation by targeting IRF1. Cell Death Dis. 2010; 1 -e94. .
- 51. Matteucci C, Minutolo A, Balestrieri E, Marino-Merlo F, Bramanti P, Garaci E, Macchi B, Mastino A. Inhibition of NF-kappaB activation sensitizes U937 cells to 3'-azido-3'-deoxythymidine induced apoptosis. Cell Death Dis. 2010; 1 -e81. .
- 52. Lei WW, Zhang KH, Pan XC, Wang DM, Hu Y, Yang YN, Song JG. Histone deacetylase 1 and 2 differentially regulate apoptosis by opposing effects on extracellular signal-regulated kinase 1/2. Cell Death Dis. 2010; 1 -e44. .
- 53. Lanzillotta A, Sarnico I, Ingrassia R, Boroni F, Branca C, Benarese M, Faraco G, Blasi F, Chiarugi A, Spano P, Pizzi M. The acetylation of RelA in Lys310 dictates the NF-kappaB-dependent response in post-ischemic injury. Cell Death Dis. 2010; 1 -e96. .
- 54. Mitchell G, Fillinger J, Sittadjody S, Avila J, Burd R, Limesand K. IGF1 activates cell cycle arrest following irradiation by reducing binding of DeltaNp63 to the p21 promoter. Cell Death Dis. 2010; 2010 -e50. .
- 55. Rello-Varona S, Kepp O, Vitale I, Michaud M, Senovilla L, Jemaa M, Joza N, Galluzzi L, Castedo M, Kroemer G. An automated fluorescence videomicroscopy assay for the detection of mitotic catastrophe. Cell Death Dis. 2010; 1 -e25. .
- 56. Knauer SK, Heinrich UR, Bier C, Habtemichael N, Docter D, Helling K, Mann WJ, Stauber RH. An otoprotective role for the apoptosis inhibitor protein survivin. Cell Death Dis. 2010; 1 -e51. .
- 57. Chan KS, Wong CH, Huang YF, Li HY. Survivin withdrawal by nuclear export failure as a physiological switch to commit cells to apoptosis. Cell Death Dis. 2010; 1 -e57. .
- 58. Engel T, Tanaka K, Jimenez-Mateos EM, Caballero-Caballero A, Prehn JH, Henshall DC. Loss of p53 results in protracted electrographic seizures and development of an aggravated epileptic phenotype following status epilepticus. Cell Death Dis. 2010; 1 -e79. .
- 59. Meley D, Spiller DG, White MR, McDowell H, Pizer B, See V. p53-mediated delayed NF-kappaB activity enhances etoposide-induced cell death in medulloblastoma. Cell Death Dis. 2010; 1 -e41. .
- 60. Gonzalez-Cano L, Herreros-Villanueva M, Fernandez-Alonso R, Ayuso-Sacido A, Meyer G, Garcia-Verdugo JM, Silva A, Marques MM, Marin MC. p73 deficiency results in impaired self renewal and premature neuronal differentiation of mouse neural progenitors independently of p53. Cell Death Dis. 2010; 1 -e109. .
- 61. Galluzzi L, Blomgren K, Kroemer G. Mitochondrial membrane permeabilization in neuronal injury. Nat Rev Neurosci. 2009; 10: 481 -94. [PubMed] .
- 62. Criollo A, Galluzzi L, Maiuri MC, Tasdemir E, Lavandero S, Kroemer G. Mitochondrial control of cell death induced by hyperosmotic stress. Apoptosis. 2007; 12: 3 -18. [PubMed] .
- 63. Ferri KF and Kroemer G. Mitochondria–the suicide organelles. Bioessays. 2001; 23: 111 -5. [PubMed] .
- 64. Hirsch T, Marzo I, Kroemer G. Role of the mitochondrial permeability transition pore in apoptosis. Biosci Rep. 1997; 17: 67 -76. [PubMed] .
- 65. Fulda S, Galluzzi L, Kroemer G. Targeting mitochondria for cancer therapy. Nat Rev Drug Discov. 2010; 9: 447 -64. [PubMed] .
- 66. Esposti MD. Bcl-2 antagonists and cancer: from the clinic, back to the bench. Cell Death Dis. 2010; 1 -e37. .
- 67. Placzek WJ, Wei J, Kitada S, Zhai D, Reed JC, Pellecchia M. A survey of the anti-apoptotic Bcl-2 subfamily expression in cancer types provides a platform to predict the efficacy of Bcl-2 antagonists in cancer therapy. Cell Death Dis. 2010; 1 -e40. .
- 68. Tischner D, Woess C, Ottina E, Villunger A. Bcl-2-regulated cell death signalling in the prevention of autoimmunity. Cell Death Dis. 2010; 1 -e48. .
- 69. McKeller MR, Herrera-Rodriguez S, Ma W, Ortiz-Quintero B, Rangel R, Cande C, Sims-Mourtada JC, Melnikova V, Kashi C, Phan LM, Chen Z, Huang P, Dunner K Jr., et al. Vital function of PRELI and essential requirement of its LEA motif. Cell Death Dis. 2010; 1 -e21. .
- 70. Sayeed A, Meng Z, Luciani G, Chen LC, Bennington JL, Dairkee SH. Negative regulation of UCP2 by TGFbeta signaling characterizes low and intermediate-grade primary breast cancer. Cell Death Dis. 2010; 1 -e53. .
- 71. Karlberg M, Ekoff M, Labi V, Strasser A, Huang D, Nilsson G. Pro-apoptotic Bax is the major and Bak an auxiliary effector in cytokine deprivation-induced mast cell apoptosis. Cell Death Dis. 2010; 1 -e43. .
- 72. Bunk EC, Konig HG, Bernas T, Engel T, Henshall DC, Kirby BP, Prehn JH. BH3-only proteins BIM and PUMA in the regulation of survival and neuronal differentiation of newly generated cells in the adult mouse hippocampus. Cell Death Dis. 2010; 1 -e15. .
- 73. Schneider-Jakob S, Corazza N, Badmann A, Sidler D, Stuber-Roos R, Keogh A, Frese S, Tschan M, Brunner T. Synergistic induction of cell death in liver tumor cells by TRAIL and chemotherapeutic drugs via the BH3-only proteins Bim and Bid. Cell Death Dis. 2010; 1 -e86. .
- 74. Sassone J, Colciago C, Marchi P, Ascardi C, Alberti L, Di Pardo A, Zippel R, Sipione S, Silani V, Ciammola A. Mutant Huntingtin induces activation of the Bcl-2/adenovirus E1B 19-kDa interacting protein (BNip3). Cell Death Dis. 2010; 1 -e7. .
- 75. Calandrella N, De Seta C, Scarsella G, Risuleo G. Carnitine reduces the lipoperoxidative damage of the membrane and apoptosis after induction of cell stress in experimental glaucoma. Cell Death Dis. 2010; 1 -e62. .
- 76. Marino ML, Fais S, Djavaheri-Mergny M, Villa A, Meschini S, Lozupone F, Venturi G, Della Mina P, Pattingre S, Rivoltini L, Codogno P, De Milito A. Proton pump inhibition induces autophagy as a survival mechanism following oxidative stress in human melanoma cells. Cell Death Dis. 2010; 1 -e87. .
- 77. Tejedo JR, Tapia-Limonchi R, Mora-Castilla S, Cahuana GM, Hmadcha A, Martin F, Bedoya FJ, Soria B. Low concentrations of nitric oxide delay the differentiation of embryonic stem cells and promote their survival. Cell Death Dis. 2010; 1 -e80. .
- 78. Dribben WH, Eisenman LN, Mennerick S. Magnesium induces neuronal apoptosis by suppressing excitability. Cell Death Dis. 2010; 1 -e63. .
- 79. Li Q, Li H, Roughton K, Wang X, Kroemer G, Blomgren K, Zhu C. Lithium reduces apoptosis and autophagy after neonatal hypoxia-ischemia. Cell Death Dis. 2010; 1 -e56. .
- 80. Flourakis M, Lehen'kyi V, Beck B, Raphael M, Vandenberghe M, Abeele FV, Roudbaraki M, Lepage G, Mauroy B, Romanin C, Shuba Y, Skryma R, Prevarskaya N. Orai1 contributes to the establishment of an apoptosis-resistant phenotype in prostate cancer cells. Cell Death Dis. 2010; 1 -e75. .
- 81. Madeo F, Tavernarakis N, Kroemer G. Can autophagy promote longevity? Nat Cell Biol. 2010; 12: 842 -6. [PubMed] .
- 82. Kroemer G, Marino G, Levine B. Autophagy and the integrated stress response. Mol Cell. 2010; 40: 280 -93. [PubMed] .
- 83. Silver N, Proctor GB, Arno M, Carpenter GH. Activation of mTOR coincides with autophagy during ligation-induced atrophy in the rat submandibular gland. Cell Death Dis. 2010; 1 -e14. .
- 84. Criollo A, Senovilla L, Authier H, Maiuri MC, Morselli E, Vitale I, Kepp O, Tasdemir E, Galluzzi L, Shen S, Tailler M, Delahaye N, Tesniere A, et al. The IKK complex contributes to the induction of autophagy. EMBO J. 2010; 29: 619 -31. [PubMed] .
- 85. Galluzzi L, Morselli E, Vicencio JM, Kepp O, Joza N, Tajeddine N, Kroemer G. Life, death and burial: multifaceted impact of autophagy. Biochem Soc Trans. 2008; 36: 786 -90. [PubMed] .
- 86. Criollo A, Senovilla L, Authier H, Maiuri MC, Morselli E, Vitale I, Kepp O, Tasdemir E, Galluzzi L, Shen S, Tailler M, Delahaye N, Tesniere A, et al. IKK connects autophagy to major stress pathways. Autophagy. 2010; 6: 189 -91. [PubMed] .
- 87. Galluzzi L, Vicencio JM, Kepp O, Tasdemir E, Maiuri MC, Kroemer G. To die or not to die: that is the autophagic question. Curr Mol Med. 2008; 8: 78 -91. [PubMed] .
- 88. Bennett HL, Fleming JT, O'Prey J, Ryan KM, Leung HY. Androgens modulate autophagy and cell death via regulation of the endoplasmic reticulum chaperone glucose-regulated protein 78/BiP in prostate cancer cells. Cell Death Dis. 2010; 1 -e72. .
- 89. Deng L, Feng J, Broaddus RR. The novel estrogen-induced gene EIG121 regulates autophagy and promotes cell survival under stress. Cell Death Dis. 2010; 1 -e32. .
- 90. Degli Esposti D, Sebagh M, Pham P, Reffas M, Pous C, Brenner C, Azoulay D, Lemoine A. Ischemic preconditioning induces autophagy and limits necrosis in human recipients of fatty liver grafts, decreasing the incidence of rejection episodes. Cell Death Dis. 2011; 2 -e111. .
- 91. Djavaheri-Mergny M, Maiuri MC, Kroemer G. Cross talk between apoptosis and autophagy by caspase-mediated cleavage of Beclin 1. Oncogene. 2010; 29: 1717 -9. [PubMed] .
- 92. Maiuri MC, Criollo A, Kroemer G. Crosstalk between apoptosis and autophagy within the Beclin 1 interactome. EMBO J. 2010; 29: 515 -6. [PubMed] .
- 93. Zamzami N, El Hamel C, Maisse C, Brenner C, Munoz-Pinedo C, Belzacq AS, Costantini P, Vieira H, Loeffler M, Molle G, Kroemer G. Bid acts on the permeability transition pore complex to induce apoptosis. Oncogene. 2000; 19: 6342 -50. [PubMed] .
- 94. Heidari N, Hicks MA, Harada H. GX15-070 (obatoclax) overcomes glucocorticoid resistance in acute lymphoblastic leukemia through induction of apoptosis and autophagy. Cell Death Dis. 2010; 1 -e76. .
- 95. McCoy F, Hurwitz J, McTavish N, Paul I, Barnes C, O'Hagan B, Odrzywol K, Murray J, Longley D, McKerr G, Fennell DA. Obatoclax induces Atg7-dependent autophagy independent of beclin-1 and BAX/BAK. Cell Death Dis. 2010; 1 -e108. .
- 96. Buttner S, Ruli D, Vogtle FN, Galluzzi L, Moitzi B, Eisenberg T, Kepp O, Habernig L, Carmona-Gutierrez D, Rockenfeller P, Laun P, Breitenbach M, Khoury C, et al. A yeast BH3-only protein mediates the mitochondrial pathway of apoptosis. EMBO J. 2011; 30: 2779 -92. [PubMed] .
- 97. Levine B, Sinha S, Kroemer G. Bcl-2 family members: dual regulators of apoptosis and autophagy. Autophagy. 2008; 4: 600 -6. [PubMed] .
- 98. Maiuri MC, Criollo A, Tasdemir E, Vicencio JM, Tajeddine N, Hickman JA, Geneste O, Kroemer G. BH3-only proteins and BH3 mimetics induce autophagy by competitively disrupting the interaction between Beclin 1 and Bcl-2/Bcl-X(L). Autophagy. 2007; 3: 374 -6. [PubMed] .
- 99. Maiuri MC, Le Toumelin G, Criollo A, Rain JC, Gautier F, Juin P, Tasdemir E, Pierron G, Troulinaki K, Tavernarakis N, Hickman JA, Geneste O, Kroemer G. Functional and physical interaction between Bcl-X(L) and a BH3-like domain in Beclin-1. EMBO J. 2007; 26: 2527 -39. [PubMed] .
- 100. Malik SA, Orhon I, Morselli E, Criollo A, Shen S, Marino G, Benyounes A, Benit P, Rustin P, Maiuri MC, Kroemer G. BH3 mimetics activate multiple pro-autophagic pathways. Oncogene. 2011; in press. .
- 101. Malik SA, Shen S, Marino G, Benyounes A, Maiuri MC, Kroemer G. BH3 mimetics reveal the network properties of autophagy-regulatory signaling cascades. Autophagy. 2011; 7: 914 -6. [PubMed] .
- 102. Ben Mosbah I, Alfany-Fernandez I, Martel C, Zaouali MA, Bintanel-Morcillo M, Rimola A, Rodes J, Brenner C, Rosello-Catafau J, Peralta C. Endoplasmic reticulum stress inhibition protects steatotic and non-steatotic livers in partial hepatectomy under ischemia-reperfusion. Cell Death Dis. 2010; 1 -e52. .
- 103. Fujita E, Dai H, Tanabe Y, Zhiling Y, Yamagata T, Miyakawa T, Tanokura M, Momoi MY, Momoi T. Autism spectrum disorder is related to endoplasmic reticulum stress induced by mutations in the synaptic cell adhesion molecule, CADM1. Cell Death Dis. 2010; 1 -e47. .
- 104. Eisenberg T, Knauer H, Schauer A, Buttner S, Ruckenstuhl C, Carmona-Gutierrez D, Ring J, Schroeder S, Magnes C, Antonacci L, Fussi H, Deszcz L, Hartl R, et al. Induction of autophagy by spermidine promotes longevity. Nat Cell Biol. 2009; 11: 1305 -14. [PubMed] .
- 105. Bjedov I, Toivonen JM, Kerr F, Slack C, Jacobson J, Foley A, Partridge L. Mechanisms of life span extension by rapamycin in the fruit fly Drosophila melanogaster. Cell Metab. 2010; 11: 35 -46. [PubMed] .
- 106. Morselli E, Marino G, Bennetzen MV, Eisenberg T, Megalou E, Schroeder S, Cabrera S, Benit P, Rustin P, Criollo A, Kepp O, Galluzzi L, Shen S, et al. Spermidine and resveratrol induce autophagy by distinct pathways converging on the acetylproteome. J Cell Biol. 2011; 192: 615 -29. [PubMed] .
- 107. Marino G, Morselli E, Bennetzen MV, Eisenberg T, Megalou E, Schroeder S, Cabrera S, Benit P, Rustin P, Criollo A, Kepp O, Galluzzi L, Shen S, et al. Longevity-relevant regulation of autophagy at the level of the acetylproteome. Autophagy. 2011; 7: 647 -9. [PubMed] .
- 108. Green DR, Galluzzi L, Kroemer G. Mitochondria and the autophagy-inflammation-cell death axis in organismal aging. Science. 2011; 333: 1109 -12. [PubMed] .
- 109. Rubinsztein DC, Marino G, Kroemer G. Autophagy and aging. Cell. 2011; 146: 682 -95. [PubMed] .
- 110. Leontieva OV, Gudkov AV, Blagosklonny MV. Weak p53 permits senescence during cell cycle arrest. Cell Cycle. 2010; 9: 4323 -7. [PubMed] .
- 111. Leontieva OV and Blagosklonny MV. DNA damaging agents and p53 do not cause senescence in quiescent cells, while consecutive re-activation of mTOR is associated with conversion to senescence. Aging (Albany NY). 2010; 2: 924 -35. [PubMed] .
- 112. Korotchkina LG, Leontieva OV, Bukreeva EI, Demidenko ZN, Gudkov AV, Blagosklonny MV. The choice between p53-induced senescence and quiescence is determined in part by the mTOR pathway. Aging (Albany NY). 2010; 2: 344 -52. [PubMed] .
- 113. Demidenko ZN, Korotchkina LG, Gudkov AV, Blagosklonny MV. Paradoxical suppression of cellular senescence by p53. Proc Natl Acad Sci U S A. 2010; 107: 9660 -4. [PubMed] .
- 114. Anisimov VN, Zabezhinski MA, Popovich IG, Piskunova TS, Semenchenko AV, Tyndyk ML, Yurova MN, Antoch MP, Blagosklonny MV. Rapamycin extends maximal lifespan in cancer-prone mice. Am J Pathol. 2010; 176: 2092 -7. [PubMed] .
- 115. Blagosklonny MV. Calorie restriction: decelerating mTOR-driven aging from cells to organisms (including humans). Cell Cycle. 2010; 9: 683 -8. [PubMed] .
- 116. Bose R, Moors M, Tofighi R, Cascante A, Hermanson O, Ceccatelli S. Glucocorticoids induce long-lasting effects in neural stem cells resulting in senescence-related alterations. Cell Death Dis. 2010; 1 -e92. .
- 117. Manning JA and Kumar S. A potential role for NEDD1 and the centrosome in senescence of mouse embryonic fibroblasts. Cell Death Dis. 2010; 1 -e35. .
- 118. Upreti M, Koonce NA, Hennings L, Chambers TC, Griffin RJ. Pegylated IFN-alpha sensitizes melanoma cells to chemotherapy and causes premature senescence in endothelial cells by IRF-1 mediated signaling. Cell Death Dis. 2010; 1 -e67. .
- 119. Wu PC, Wang Q, Dong ZM, Chu E, Roberson RS, Ivanova IC, Wu DY. Expression of coxsackie and adenovirus receptor distinguishes transitional cancer states in therapy-induced cellular senescence. Cell Death Dis. 2010; 1 -e70. .
- 120. Blagosklonny MV. Increasing healthy lifespan by suppressing aging in our lifetime: preliminary proposal. Cell Cycle. 2010; 9: 4788 -94. [PubMed] .
- 121. Blagosklonny MV. Why human lifespan is rapidly increasing: solving “longevity riddle” with “revealed-slow-aging” hypothesis. Aging (Albany NY). 2010; 2: 177 -82. [PubMed] .
- 122. Blagosklonny MV. Why men age faster but reproduce longer than women: mTOR and evolutionary perspectives. Aging (Albany NY). 2010; 2: 265 -73. [PubMed] .
- 123. Blagosklonny MV. Why the disposable soma theory cannot explain why women live longer and why we age. Aging (Albany NY). 2010; 2: 884 -7. [PubMed] .
- 124. Maiuri MC, Galluzzi L, Morselli E, Kepp O, Malik SA, Kroemer G. Autophagy regulation by p53. Curr Opin Cell Biol. 2010; 22: 181 -5. [PubMed] .
- 125. Morselli E, Galluzzi L, Kepp O, Marino G, Michaud M, Vitale I, Maiuri MC, Kroemer G. Oncosuppressive functions of autophagy. Antioxid Redox Signal. 2011; 14: 2251 -69. [PubMed] .
- 126. Maiuri MC, Tasdemir E, Criollo A, Morselli E, Vicencio JM, Carnuccio R, Kroemer G. Control of autophagy by oncogenes and tumor suppressor genes. Cell Death Differ. 2009; 16: 87 -93. [PubMed] .
- 127. Rabinowitz JD and White E. Autophagy and metabolism. Science. 2010; 330: 1344 -8. [PubMed] .
- 128. Castedo M, Perfettini JL, Roumier T, Valent A, Raslova H, Yakushijin K, Horne D, Feunteun J, Lenoir G, Medema R, Vainchenker W, Kroemer G. Mitotic catastrophe constitutes a special case of apoptosis whose suppression entails aneuploidy. Oncogene. 2004; 23: 4362 -70. [PubMed] .
- 129. Vitale I, Galluzzi L, Castedo M, Kroemer G. Mitotic catastrophe: a mechanism for avoiding genomic instability. Nat Rev Mol Cell Biol. 2011; 12: 385 -92. [PubMed] .
- 130. Vitale I, Galluzzi L, Senovilla L, Criollo A, Jemaa M, Castedo M, Kroemer G. Illicit survival of cancer cells during polyploidization and depolyploidization. Cell Death Differ. 2011; 18: 1403 -13. [PubMed] .
- 131. Yacoubian TA, Slone SR, Harrington AJ, Hamamichi S, Schieltz JM, Caldwell KA, Caldwell GA, Standaert DG. Differential neuroprotective effects of 14-3-3 proteins in models of Parkinson's disease. Cell Death Dis. 2010; 1 -e2. .
- 132. Rodriguez JJ, Witton J, Olabarria M, Noristani HN, Verkhratsky A. Increase in the density of resting microglia precedes neuritic plaque formation and microglial activation in a transgenic model of Alzheimer's disease. Cell Death Dis. 2010; 1 -e1. .
- 133. Sikkink LA and Ramirez-Alvarado M. Cytotoxicity of amyloidogenic immunoglobulin light chains in cell culture. Cell Death Dis. 2010; 1 -e98. .
- 134. Ciavardelli D, Silvestri E, Viscovo AD, Bomba M, Gregorio DD, Moreno M, Ilio CD, Goglia F, Canzoniero LM, Sensi SL. Alterations of brain and cerebellar proteomes linked to Abeta and tau pathology in a female triple-transgenic murine model of Alzheimer's disease. Cell Death Dis. 2010; 1 -e90. .