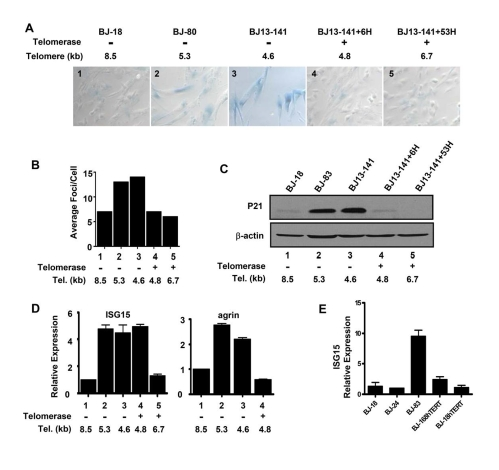
Figure 2.Replicative senescence and DNA damage signaling independent up-regulation of ISG15 expression in cells with short telomeres. (A)
Short and long-term expression of hTERT rescued cells from replicative
senescence. BJ cells with short telomeres (BJ-80 and BJ13-141) exhibited
significant increases in the number of SA-β-Gal positive cells;
whereas, the cells with long telomeres (BJ-18) did not show SA-β-Gal
staining. Exogenous telomerase rapidly eliminated senescent cells
(BJ13-141+6H, when only the shortest telomeres had been lengthened) as well
as after bulk telomere elongation had occurred (BJ13-141+53H). Rare fields
with an SA-β-Gal staining positive cell were selected for the last two
images to validate the staining procedure. The number in each image is a
key to the cell lines used in B-D. (B) γ-H2AX staining shows
that exogenous hTERT rapidly eliminates DNA damage signalling due to short
telomeres. Approximately 500 nuclei of each cell line were analyzed using
Metasystems software (Metasystems, Germany). (C) Western blot shows
that p21, a transcriptional target of DNA damage-induced p53 signaling,
rapidly disappeared following the introduction of telomerase to elongate
the shortest telomeres. (D) Q-PCR showing that ISG15
expression remained high in BJ cells rescued from replicative
senescence/DNA damage signaling after only a few doublings in the presence
of exogenous telomerase when telomeres were still short (ISG15, column 4),
while elongation of the telomeres after 53 doublings led to decreased
expression (ISG15, column 5). In contrast, elimi-nating replicative senescence/DNA damage following a short exposure
to telomerase caused a decrease in the expression of agrin (agrin, column
4). Agrin thus did not meet our criteria for telomere length regulation,
since its increase in old cells (agrin, columns 2&3) is secondary to
senescence and/or DNA damage (column 4). (E) BJ cells
overexpressing hTERT and having long telomeres express low levels of ISG15.