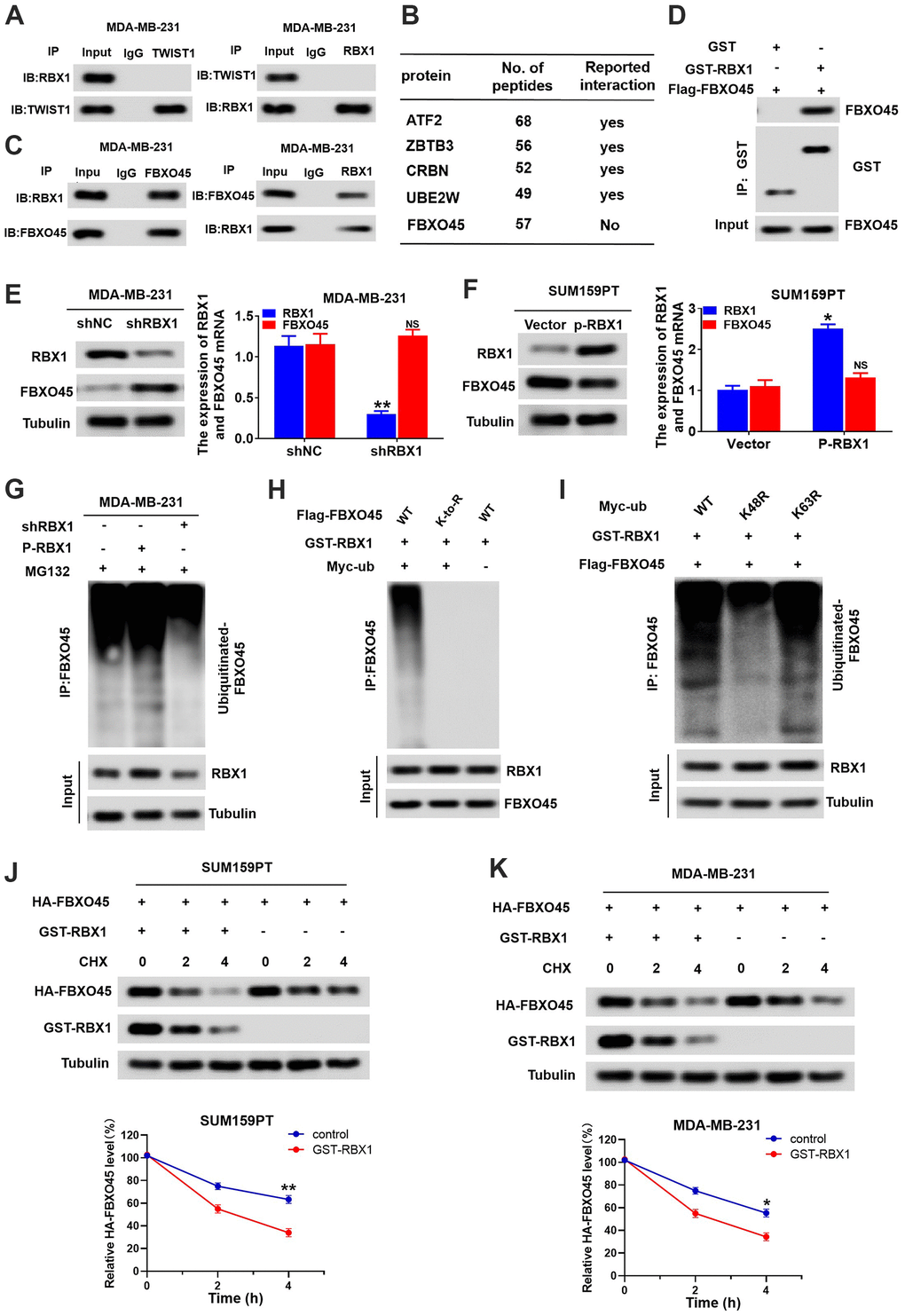
Figure 6.RBX1 promotes FBXO45 ubiquitination and degradation in TNBC cells. (A) Co-IP reveals no direct binding of endogenous TWIST1and RBX1. (B) Partial list of proteins related to RBX1, determined with immunoprecipitation-mass spectrometry. (C) Co-IP reflected direct binding of endogenous FBXO45 and RBX1 in the MDA-MB-231 cells. (D) GST pull-down analysis displaying direct binding of FBXO45 and RBX1. (E) qRT-PCR and western blotting exhibiting the FBXO45 and RBX1 expression levels in the RBX1-silencing MDA-MB-231 cells. **P < 0.01. (F) The qRT-PCR and western blotting revealing the FBXO45 and RBX1 expression levels in SUM159PT cells stably transfected by plasmid with overexpression of RBX1. **P < 0.01. (G) RBX1 exogenous expression or knockdown changed the FBXO45 ubiquitination. The proteasome suppressor MG132 was used to treat the cells from each group. Cell lysates were generated and next immunoprecipitated by an anti-FBXO45 antibody. Western blotting was applied for measuring the levels of ubiquitin-linked FBBXO45 utilizing anti-UB antibody. (H) Ubiquitination of wild-type FBBXO45 or K-to-R mutants (mutations in all Lys position of the FBBXO45 gene). (I) FBXO45 ubiquitination in the HEK293 cells was detected. (J, K) TNBC cells were transfected by plasmids encoding HA-FBXO45, without or with p-RBX1 plasmid. Cells were next exposed to 20μmol/L of cyclohexanone (CHX) at the given times, and the anti-HA antibody was utilized to determine FBXO45 degradation. *P < 0.05, **P < 0.01.